Lecture Notes on Human Respiratory System Physiology
VerifiedAdded on 2023/02/03
|33
|9430
|43
AI Summary
This document provides comprehensive lecture notes on the mechanics of breathing, regulation and control of breathing, ventilation, lung volumes and pulmonary function tests, diffusion, perfusion, gas transport to the periphery, acid-base regulation, respiratory system under stress, and more.
Contribute Materials
Your contribution can guide someone’s learning journey. Share your
documents today.
1
LECTURE NOTES ON
HUMAN RESPIRATORY SYSTEM PHYSIOLOGY
(Dr. GÜL ERDEMLI)
CONTENTS
1. MECHANICS OF BREATHING:
2. REGULATION AND CONTROL OF BREATHING:
3. VENTILATION
4. LUNG VOLUMES AND PULMONARY FUNCTION TESTS
5. DIFFUSION
6. PERFUSION
7. GAS TRANSPORT TO THE PERIPHERY
8. ACID-BASE REGULATION
9. RESPIRATORY SYSTEM UNDER STRESS
10. RECOMMENDED FURTHER READING:
11. SELF ASSESSMENT
LECTURE NOTES ON
HUMAN RESPIRATORY SYSTEM PHYSIOLOGY
(Dr. GÜL ERDEMLI)
CONTENTS
1. MECHANICS OF BREATHING:
2. REGULATION AND CONTROL OF BREATHING:
3. VENTILATION
4. LUNG VOLUMES AND PULMONARY FUNCTION TESTS
5. DIFFUSION
6. PERFUSION
7. GAS TRANSPORT TO THE PERIPHERY
8. ACID-BASE REGULATION
9. RESPIRATORY SYSTEM UNDER STRESS
10. RECOMMENDED FURTHER READING:
11. SELF ASSESSMENT
Secure Best Marks with AI Grader
Need help grading? Try our AI Grader for instant feedback on your assignments.
2
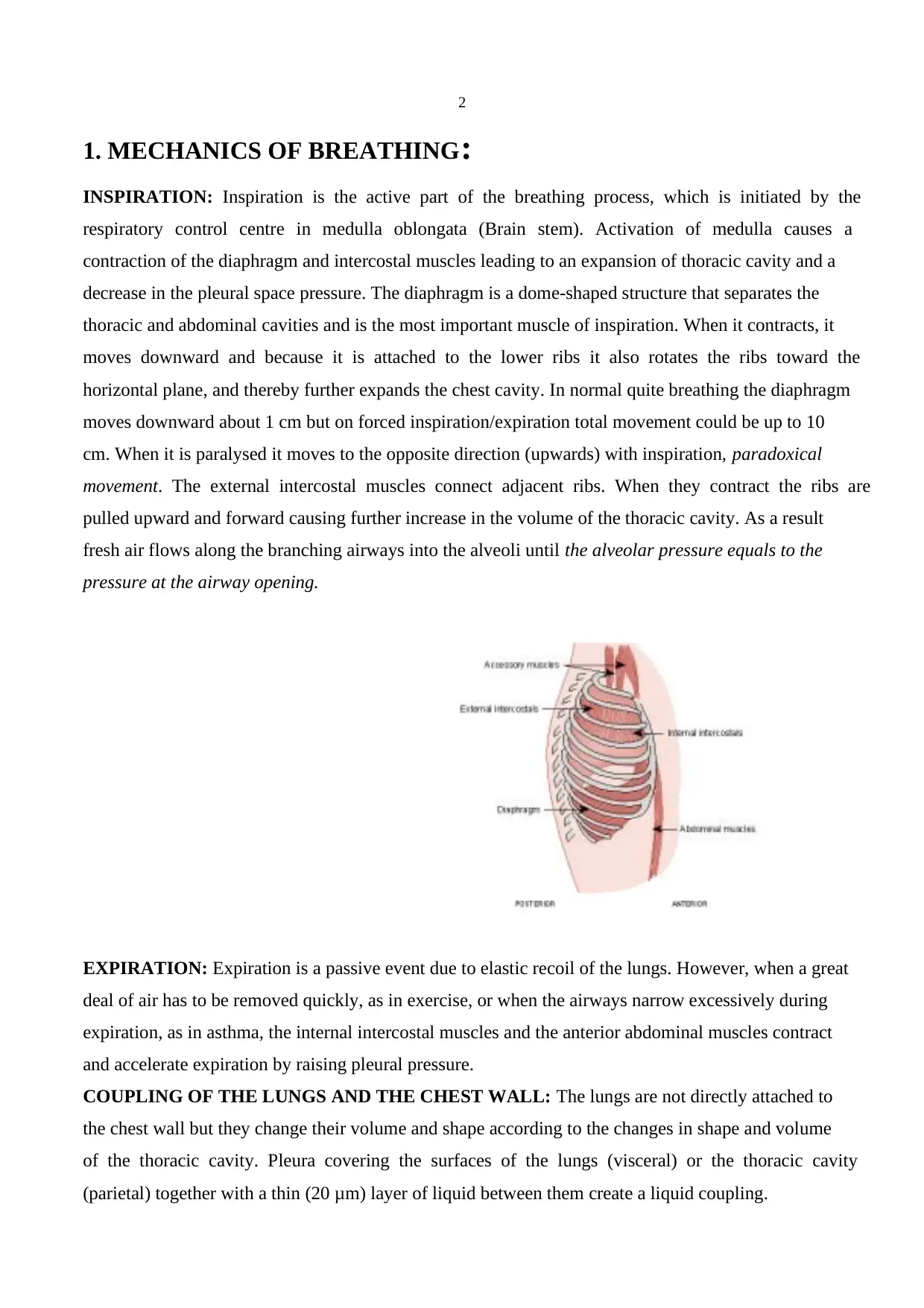
1. MECHANICS OF BREATHING:
INSPIRATION: Inspiration is the active part of the breathing process, which is initiated by the
respiratory control centre in medulla oblongata (Brain stem). Activation of medulla causes a
contraction of the diaphragm and intercostal muscles leading to an expansion of thoracic cavity and a
decrease in the pleural space pressure. The diaphragm is a dome-shaped structure that separates the
thoracic and abdominal cavities and is the most important muscle of inspiration. When it contracts, it
moves downward and because it is attached to the lower ribs it also rotates the ribs toward the
horizontal plane, and thereby further expands the chest cavity. In normal quite breathing the diaphragm
moves downward about 1 cm but on forced inspiration/expiration total movement could be up to 10
cm. When it is paralysed it moves to the opposite direction (upwards) with inspiration, paradoxical
movement. The external intercostal muscles connect adjacent ribs. When they contract the ribs are
pulled upward and forward causing further increase in the volume of the thoracic cavity. As a result
fresh air flows along the branching airways into the alveoli until the alveolar pressure equals to the
pressure at the airway opening.
EXPIRATION: Expiration is a passive event due to elastic recoil of the lungs. However, when a great
deal of air has to be removed quickly, as in exercise, or when the airways narrow excessively during
expiration, as in asthma, the internal intercostal muscles and the anterior abdominal muscles contract
and accelerate expiration by raising pleural pressure.
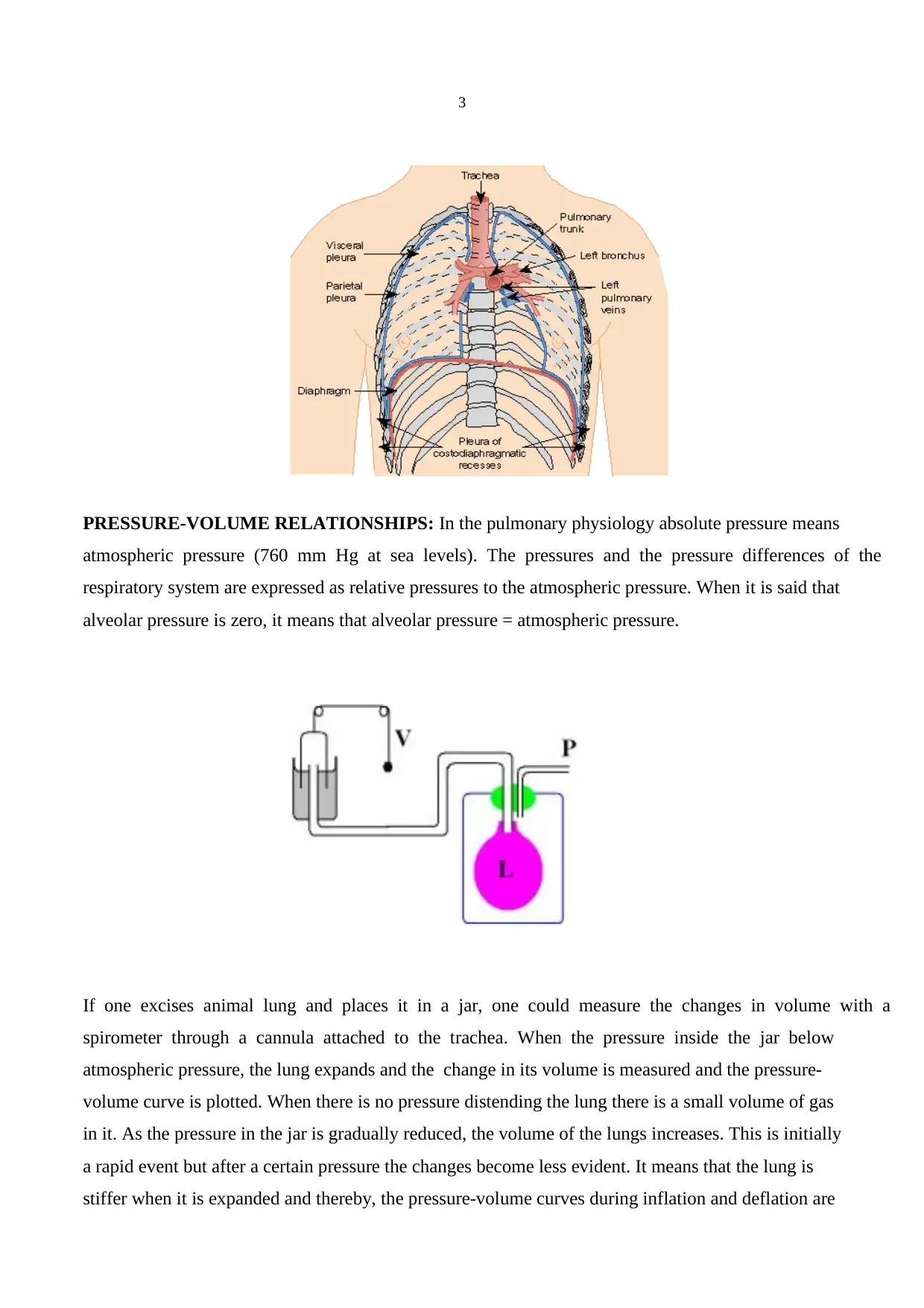
COUPLING OF THE LUNGS AND THE CHEST WALL: The lungs are not directly attached to
the chest wall but they change their volume and shape according to the changes in shape and volume
of the thoracic cavity. Pleura covering the surfaces of the lungs (visceral) or the thoracic cavity
(parietal) together with a thin (20 μm) layer of liquid between them create a liquid coupling.
1. MECHANICS OF BREATHING:
INSPIRATION: Inspiration is the active part of the breathing process, which is initiated by the
respiratory control centre in medulla oblongata (Brain stem). Activation of medulla causes a
contraction of the diaphragm and intercostal muscles leading to an expansion of thoracic cavity and a
decrease in the pleural space pressure. The diaphragm is a dome-shaped structure that separates the
thoracic and abdominal cavities and is the most important muscle of inspiration. When it contracts, it
moves downward and because it is attached to the lower ribs it also rotates the ribs toward the
horizontal plane, and thereby further expands the chest cavity. In normal quite breathing the diaphragm
moves downward about 1 cm but on forced inspiration/expiration total movement could be up to 10
cm. When it is paralysed it moves to the opposite direction (upwards) with inspiration, paradoxical
movement. The external intercostal muscles connect adjacent ribs. When they contract the ribs are
pulled upward and forward causing further increase in the volume of the thoracic cavity. As a result
fresh air flows along the branching airways into the alveoli until the alveolar pressure equals to the
pressure at the airway opening.
EXPIRATION: Expiration is a passive event due to elastic recoil of the lungs. However, when a great
deal of air has to be removed quickly, as in exercise, or when the airways narrow excessively during
expiration, as in asthma, the internal intercostal muscles and the anterior abdominal muscles contract
and accelerate expiration by raising pleural pressure.
COUPLING OF THE LUNGS AND THE CHEST WALL: The lungs are not directly attached to
the chest wall but they change their volume and shape according to the changes in shape and volume
of the thoracic cavity. Pleura covering the surfaces of the lungs (visceral) or the thoracic cavity
(parietal) together with a thin (20 μm) layer of liquid between them create a liquid coupling.
3
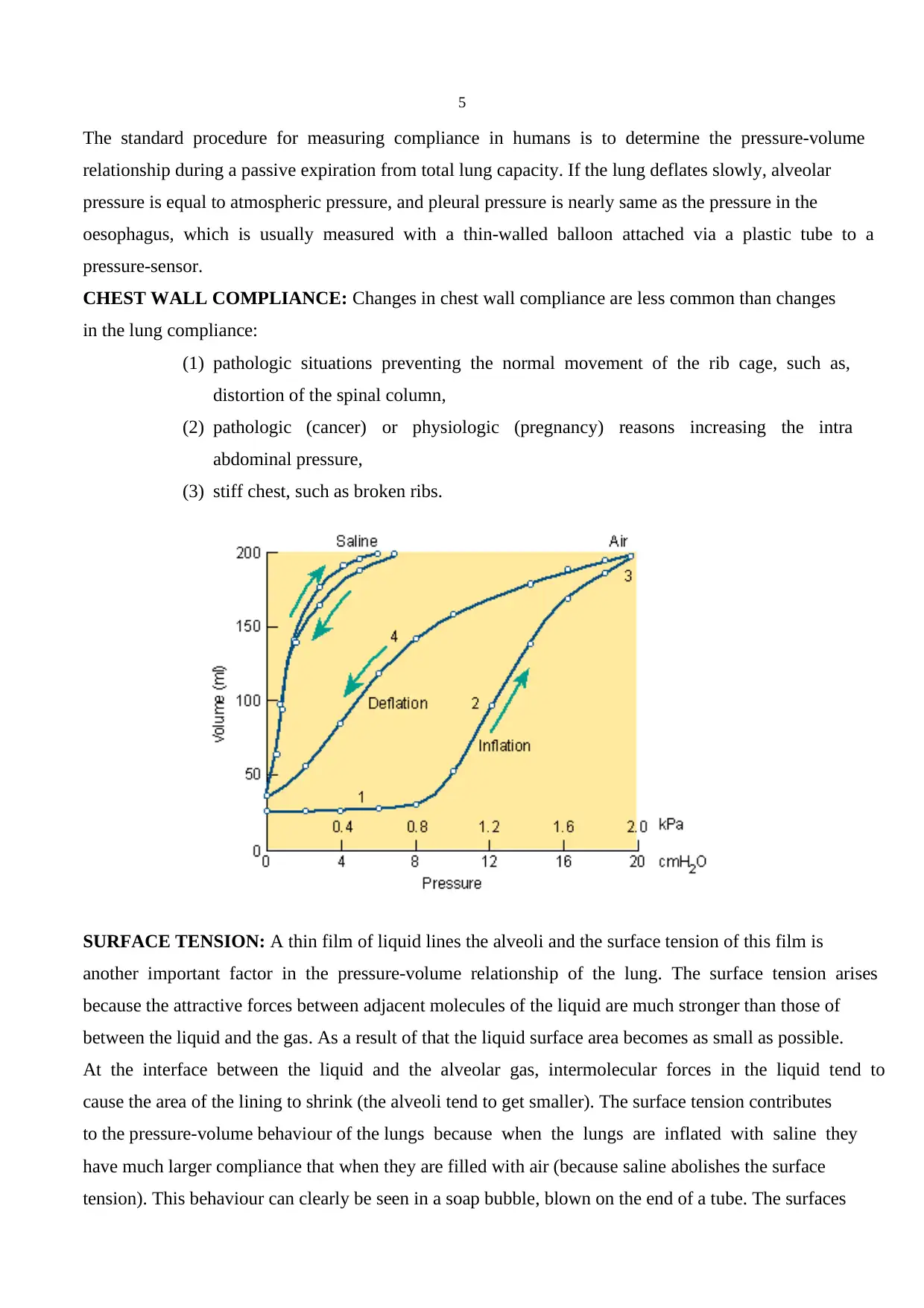
PRESSURE-VOLUME RELATIONSHIPS: In the pulmonary physiology absolute pressure means
atmospheric pressure (760 mm Hg at sea levels). The pressures and the pressure differences of the
respiratory system are expressed as relative pressures to the atmospheric pressure. When it is said that
alveolar pressure is zero, it means that alveolar pressure = atmospheric pressure.
If one excises animal lung and places it in a jar, one could measure the changes in volume with a
spirometer through a cannula attached to the trachea. When the pressure inside the jar below
atmospheric pressure, the lung expands and the change in its volume is measured and the pressure-
volume curve is plotted. When there is no pressure distending the lung there is a small volume of gas
in it. As the pressure in the jar is gradually reduced, the volume of the lungs increases. This is initially
a rapid event but after a certain pressure the changes become less evident. It means that the lung is
stiffer when it is expanded and thereby, the pressure-volume curves during inflation and deflation are
PRESSURE-VOLUME RELATIONSHIPS: In the pulmonary physiology absolute pressure means
atmospheric pressure (760 mm Hg at sea levels). The pressures and the pressure differences of the
respiratory system are expressed as relative pressures to the atmospheric pressure. When it is said that
alveolar pressure is zero, it means that alveolar pressure = atmospheric pressure.
If one excises animal lung and places it in a jar, one could measure the changes in volume with a
spirometer through a cannula attached to the trachea. When the pressure inside the jar below
atmospheric pressure, the lung expands and the change in its volume is measured and the pressure-
volume curve is plotted. When there is no pressure distending the lung there is a small volume of gas
in it. As the pressure in the jar is gradually reduced, the volume of the lungs increases. This is initially
a rapid event but after a certain pressure the changes become less evident. It means that the lung is
stiffer when it is expanded and thereby, the pressure-volume curves during inflation and deflation are
4
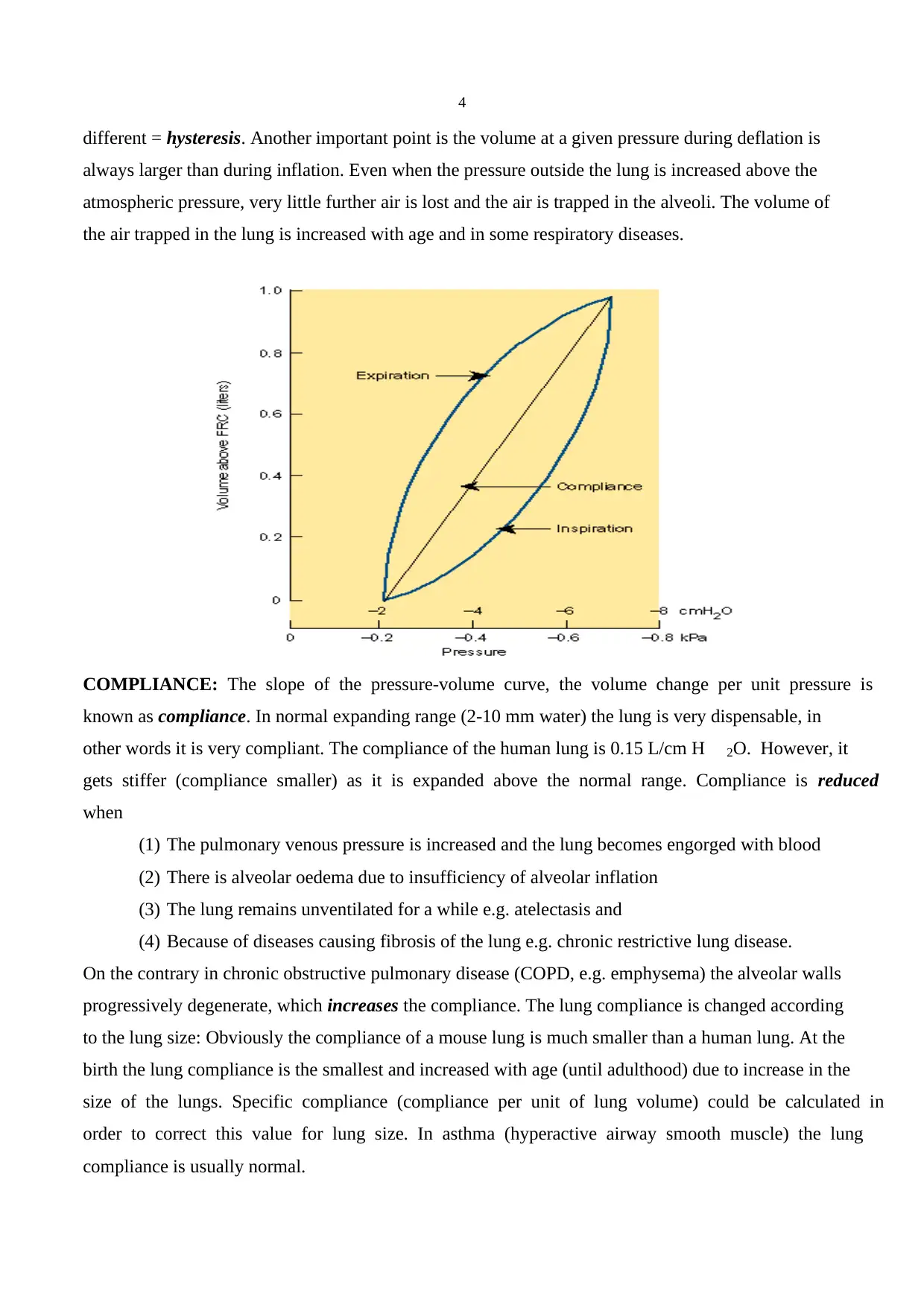
different = hysteresis. Another important point is the volume at a given pressure during deflation is
always larger than during inflation. Even when the pressure outside the lung is increased above the
atmospheric pressure, very little further air is lost and the air is trapped in the alveoli. The volume of
the air trapped in the lung is increased with age and in some respiratory diseases.
COMPLIANCE: The slope of the pressure-volume curve, the volume change per unit pressure is
known as compliance. In normal expanding range (2-10 mm water) the lung is very dispensable, in
other words it is very compliant. The compliance of the human lung is 0.15 L/cm H 2O. However, it
gets stiffer (compliance smaller) as it is expanded above the normal range. Compliance is reduced
when
(1) The pulmonary venous pressure is increased and the lung becomes engorged with blood
(2) There is alveolar oedema due to insufficiency of alveolar inflation
(3) The lung remains unventilated for a while e.g. atelectasis and
(4) Because of diseases causing fibrosis of the lung e.g. chronic restrictive lung disease.
On the contrary in chronic obstructive pulmonary disease (COPD, e.g. emphysema) the alveolar walls
progressively degenerate, which increases the compliance. The lung compliance is changed according
to the lung size: Obviously the compliance of a mouse lung is much smaller than a human lung. At the
birth the lung compliance is the smallest and increased with age (until adulthood) due to increase in the
size of the lungs. Specific compliance (compliance per unit of lung volume) could be calculated in
order to correct this value for lung size. In asthma (hyperactive airway smooth muscle) the lung
compliance is usually normal.
different = hysteresis. Another important point is the volume at a given pressure during deflation is
always larger than during inflation. Even when the pressure outside the lung is increased above the
atmospheric pressure, very little further air is lost and the air is trapped in the alveoli. The volume of
the air trapped in the lung is increased with age and in some respiratory diseases.
COMPLIANCE: The slope of the pressure-volume curve, the volume change per unit pressure is
known as compliance. In normal expanding range (2-10 mm water) the lung is very dispensable, in
other words it is very compliant. The compliance of the human lung is 0.15 L/cm H 2O. However, it
gets stiffer (compliance smaller) as it is expanded above the normal range. Compliance is reduced
when
(1) The pulmonary venous pressure is increased and the lung becomes engorged with blood
(2) There is alveolar oedema due to insufficiency of alveolar inflation
(3) The lung remains unventilated for a while e.g. atelectasis and
(4) Because of diseases causing fibrosis of the lung e.g. chronic restrictive lung disease.
On the contrary in chronic obstructive pulmonary disease (COPD, e.g. emphysema) the alveolar walls
progressively degenerate, which increases the compliance. The lung compliance is changed according
to the lung size: Obviously the compliance of a mouse lung is much smaller than a human lung. At the
birth the lung compliance is the smallest and increased with age (until adulthood) due to increase in the
size of the lungs. Specific compliance (compliance per unit of lung volume) could be calculated in
order to correct this value for lung size. In asthma (hyperactive airway smooth muscle) the lung
compliance is usually normal.
Secure Best Marks with AI Grader
Need help grading? Try our AI Grader for instant feedback on your assignments.
5
The standard procedure for measuring compliance in humans is to determine the pressure-volume
relationship during a passive expiration from total lung capacity. If the lung deflates slowly, alveolar
pressure is equal to atmospheric pressure, and pleural pressure is nearly same as the pressure in the
oesophagus, which is usually measured with a thin-walled balloon attached via a plastic tube to a
pressure-sensor.
CHEST WALL COMPLIANCE: Changes in chest wall compliance are less common than changes
in the lung compliance:
(1) pathologic situations preventing the normal movement of the rib cage, such as,
distortion of the spinal column,
(2) pathologic (cancer) or physiologic (pregnancy) reasons increasing the intra
abdominal pressure,
(3) stiff chest, such as broken ribs.
SURFACE TENSION: A thin film of liquid lines the alveoli and the surface tension of this film is
another important factor in the pressure-volume relationship of the lung. The surface tension arises
because the attractive forces between adjacent molecules of the liquid are much stronger than those of
between the liquid and the gas. As a result of that the liquid surface area becomes as small as possible.
At the interface between the liquid and the alveolar gas, intermolecular forces in the liquid tend to
cause the area of the lining to shrink (the alveoli tend to get smaller). The surface tension contributes
to the pressure-volume behaviour of the lungs because when the lungs are inflated with saline they
have much larger compliance that when they are filled with air (because saline abolishes the surface
tension). This behaviour can clearly be seen in a soap bubble, blown on the end of a tube. The surfaces
The standard procedure for measuring compliance in humans is to determine the pressure-volume
relationship during a passive expiration from total lung capacity. If the lung deflates slowly, alveolar
pressure is equal to atmospheric pressure, and pleural pressure is nearly same as the pressure in the
oesophagus, which is usually measured with a thin-walled balloon attached via a plastic tube to a
pressure-sensor.
CHEST WALL COMPLIANCE: Changes in chest wall compliance are less common than changes
in the lung compliance:
(1) pathologic situations preventing the normal movement of the rib cage, such as,
distortion of the spinal column,
(2) pathologic (cancer) or physiologic (pregnancy) reasons increasing the intra
abdominal pressure,
(3) stiff chest, such as broken ribs.
SURFACE TENSION: A thin film of liquid lines the alveoli and the surface tension of this film is
another important factor in the pressure-volume relationship of the lung. The surface tension arises
because the attractive forces between adjacent molecules of the liquid are much stronger than those of
between the liquid and the gas. As a result of that the liquid surface area becomes as small as possible.
At the interface between the liquid and the alveolar gas, intermolecular forces in the liquid tend to
cause the area of the lining to shrink (the alveoli tend to get smaller). The surface tension contributes
to the pressure-volume behaviour of the lungs because when the lungs are inflated with saline they
have much larger compliance that when they are filled with air (because saline abolishes the surface
tension). This behaviour can clearly be seen in a soap bubble, blown on the end of a tube. The surfaces

6
of the bubble contract as much as possible and form the smallest possible surface area, a sphere. This
generates a pressure predicted from Laplace’s law:
Pressure =(4 x surface tension) / radius
The surface tension contributes a large part of the static recoil force of the lung (expiration). The
surface tension changes with the surface area: The larger the area the smaller the surface tension gets.
The most important component of this liquid film is surfactant. It is produced by type 2 alveolar
epithelial cells and its major constituent is dipalmitoyl phosphotidylcholine (DPPC), a phospholipid
with detergent properties. The phospholipid DPPC is synthesised in the lung from fatty acids that are
either extracted from the blood or are themselves synthesised in the lung. Synthesis is fast and there is
a rapid turnover of surfactant. If the blood flow to a region of lung is restricted due to an embolus the
surfactant may be depleted in the effected area. Surfactant synthesis starts relatively late in foetal life
and premature babies without adequate amount of surfactant develop respiratory distress which could
be life threatening.
What are the advantages of having surfactant and the low surface tension?
1. It increases the compliance of the lung
2. It reduces the work of expanding of the lung with each breath
3. It stabilises the alveoli (thus the smaller alveoli do not collapse at the end-expiration)
4. It keeps the alveoli dry (as the surface tension tends to collapse alveoli, it also tends to suck fluid
into the alveolar space from capillaries).
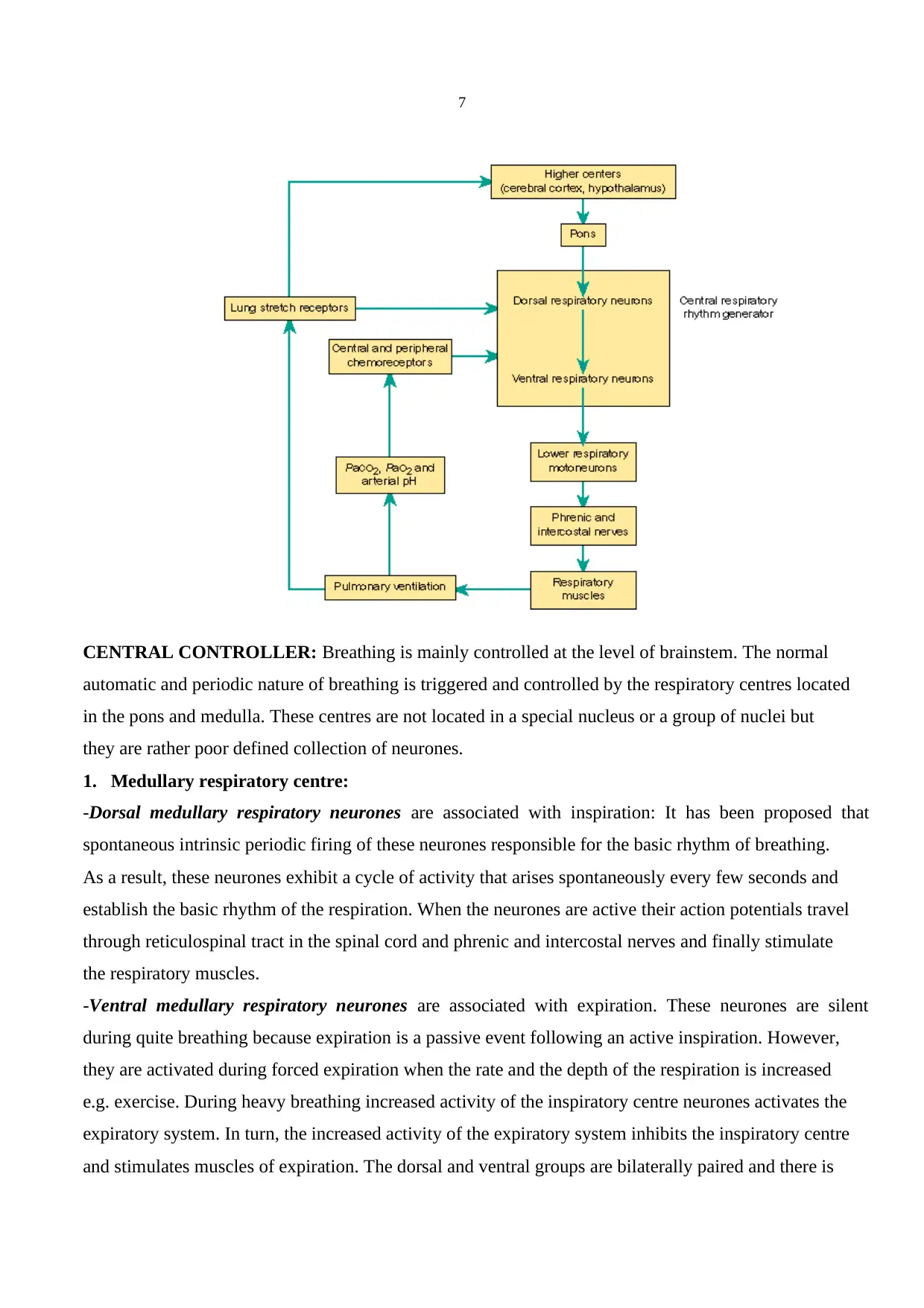
2. REGULATION AND CONTROL OF BREATHING:
In order to maintain normal levels of partial oxygen and carbon dioxide pressure both the depth and
rate of breathing are precisely regulated. Basic elements of the respiratory control system are (1)
strategically placed sensors (2) central controller (3) respiratory muscles.
of the bubble contract as much as possible and form the smallest possible surface area, a sphere. This
generates a pressure predicted from Laplace’s law:
Pressure =(4 x surface tension) / radius
The surface tension contributes a large part of the static recoil force of the lung (expiration). The
surface tension changes with the surface area: The larger the area the smaller the surface tension gets.
The most important component of this liquid film is surfactant. It is produced by type 2 alveolar
epithelial cells and its major constituent is dipalmitoyl phosphotidylcholine (DPPC), a phospholipid
with detergent properties. The phospholipid DPPC is synthesised in the lung from fatty acids that are
either extracted from the blood or are themselves synthesised in the lung. Synthesis is fast and there is
a rapid turnover of surfactant. If the blood flow to a region of lung is restricted due to an embolus the
surfactant may be depleted in the effected area. Surfactant synthesis starts relatively late in foetal life
and premature babies without adequate amount of surfactant develop respiratory distress which could
be life threatening.
What are the advantages of having surfactant and the low surface tension?
1. It increases the compliance of the lung
2. It reduces the work of expanding of the lung with each breath
3. It stabilises the alveoli (thus the smaller alveoli do not collapse at the end-expiration)
4. It keeps the alveoli dry (as the surface tension tends to collapse alveoli, it also tends to suck fluid
into the alveolar space from capillaries).
2. REGULATION AND CONTROL OF BREATHING:
In order to maintain normal levels of partial oxygen and carbon dioxide pressure both the depth and
rate of breathing are precisely regulated. Basic elements of the respiratory control system are (1)
strategically placed sensors (2) central controller (3) respiratory muscles.
7
CENTRAL CONTROLLER: Breathing is mainly controlled at the level of brainstem. The normal
automatic and periodic nature of breathing is triggered and controlled by the respiratory centres located
in the pons and medulla. These centres are not located in a special nucleus or a group of nuclei but
they are rather poor defined collection of neurones.
1. Medullary respiratory centre:
-Dorsal medullary respiratory neurones are associated with inspiration: It has been proposed that
spontaneous intrinsic periodic firing of these neurones responsible for the basic rhythm of breathing.
As a result, these neurones exhibit a cycle of activity that arises spontaneously every few seconds and
establish the basic rhythm of the respiration. When the neurones are active their action potentials travel
through reticulospinal tract in the spinal cord and phrenic and intercostal nerves and finally stimulate
the respiratory muscles.
-Ventral medullary respiratory neurones are associated with expiration. These neurones are silent
during quite breathing because expiration is a passive event following an active inspiration. However,
they are activated during forced expiration when the rate and the depth of the respiration is increased
e.g. exercise. During heavy breathing increased activity of the inspiratory centre neurones activates the
expiratory system. In turn, the increased activity of the expiratory system inhibits the inspiratory centre
and stimulates muscles of expiration. The dorsal and ventral groups are bilaterally paired and there is
CENTRAL CONTROLLER: Breathing is mainly controlled at the level of brainstem. The normal
automatic and periodic nature of breathing is triggered and controlled by the respiratory centres located
in the pons and medulla. These centres are not located in a special nucleus or a group of nuclei but
they are rather poor defined collection of neurones.
1. Medullary respiratory centre:
-Dorsal medullary respiratory neurones are associated with inspiration: It has been proposed that
spontaneous intrinsic periodic firing of these neurones responsible for the basic rhythm of breathing.
As a result, these neurones exhibit a cycle of activity that arises spontaneously every few seconds and
establish the basic rhythm of the respiration. When the neurones are active their action potentials travel
through reticulospinal tract in the spinal cord and phrenic and intercostal nerves and finally stimulate
the respiratory muscles.
-Ventral medullary respiratory neurones are associated with expiration. These neurones are silent
during quite breathing because expiration is a passive event following an active inspiration. However,
they are activated during forced expiration when the rate and the depth of the respiration is increased
e.g. exercise. During heavy breathing increased activity of the inspiratory centre neurones activates the
expiratory system. In turn, the increased activity of the expiratory system inhibits the inspiratory centre
and stimulates muscles of expiration. The dorsal and ventral groups are bilaterally paired and there is
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

8
cross communication between them. As a consequence they behave in synchrony and the respiratory
movements are symmetric.
2.Apneustic Centre: It is located in the lower pons. Exact role of this centre in the normal breathing is
not known. Lesions covering this area in the pons cause a pathologic respiratory rhythm with increased
apnoea frequency. What is known is nerve impulses from the apneustic centre stimulate the inspiratory
centre and without constant influence of this centre respiration becomes shallow and irregular.
3.Pneumotaxic centre: It is located in the upper pons. This centre is a group of neurones that have an
inhibitory effect on the both inspiratory and apneustic centres. It is probably responsible for the
termination of inspiration by inhibiting the activity of the dorsal medullar neurones. It primarily
regulates the volume and secondarily the rate of the respiration. Because in the lesions of this area
normal respiration is protected it is generally believed that upper pons is responsible for the fine-tuning
of the respiratory rhythm. Hypoactivation of this centre causes prolonged deep inspirations and brief,
limited expirations by allowing the inspiration centre remain active longer than normal.
Hyperactivation of this centre on the other hand results in shallow inspirations. The apneustic and
pneumotaxic centres function in co-ordination in order to provide a rhythmic respiratory cycle:
Activation of the inspiratory centre stimulates the muscles of inspiration and also the pneumotaxic
centre. Then the pneumotaxic centre inhibits both the apneustic and the inspiratory centres resulting in
initiation of expiration. Spontaneous activity of the neurones in the inspiratory centre starts another
similar cycle again.
Breathing in some extent is also controlled consciously from higher brain centres (e.g. cerebral cortex).
This control is required when we talk, cough and vomit. It is also possible voluntarily change the rate
of the breathing. Hyperventilation can decrease blood partial carbon dioxide pressure (PCO 2) due to
loss of CO 2 resulting in peripheral vasodilatation and decrease in blood pressure. One can also stop
breathing voluntarily. That results in an increase in arterial partial oxygen pressure (PO2), which
produces an urge to breathe. When eventually PCO2 reaches the high enough level it overrides the
conscious influences from the cortex and stimulates the inspiratory system. If one holds his breath long
enough to decrease PO2 to a very low level one may loose his consciousness. In an unconscious person
automatic control of the respiration takes over and the normal breathing resumes.
Other parts of the brain (limbic system, hypothalamus) can also alter the breathing pattern e.g.
affective states, strong emotions such as rage and fear. In addition, stimulation of touch, thermal and
pain receptors can also stimulate the respiratory system.
cross communication between them. As a consequence they behave in synchrony and the respiratory
movements are symmetric.
2.Apneustic Centre: It is located in the lower pons. Exact role of this centre in the normal breathing is
not known. Lesions covering this area in the pons cause a pathologic respiratory rhythm with increased
apnoea frequency. What is known is nerve impulses from the apneustic centre stimulate the inspiratory
centre and without constant influence of this centre respiration becomes shallow and irregular.
3.Pneumotaxic centre: It is located in the upper pons. This centre is a group of neurones that have an
inhibitory effect on the both inspiratory and apneustic centres. It is probably responsible for the
termination of inspiration by inhibiting the activity of the dorsal medullar neurones. It primarily
regulates the volume and secondarily the rate of the respiration. Because in the lesions of this area
normal respiration is protected it is generally believed that upper pons is responsible for the fine-tuning
of the respiratory rhythm. Hypoactivation of this centre causes prolonged deep inspirations and brief,
limited expirations by allowing the inspiration centre remain active longer than normal.
Hyperactivation of this centre on the other hand results in shallow inspirations. The apneustic and
pneumotaxic centres function in co-ordination in order to provide a rhythmic respiratory cycle:
Activation of the inspiratory centre stimulates the muscles of inspiration and also the pneumotaxic
centre. Then the pneumotaxic centre inhibits both the apneustic and the inspiratory centres resulting in
initiation of expiration. Spontaneous activity of the neurones in the inspiratory centre starts another
similar cycle again.
Breathing in some extent is also controlled consciously from higher brain centres (e.g. cerebral cortex).
This control is required when we talk, cough and vomit. It is also possible voluntarily change the rate
of the breathing. Hyperventilation can decrease blood partial carbon dioxide pressure (PCO 2) due to
loss of CO 2 resulting in peripheral vasodilatation and decrease in blood pressure. One can also stop
breathing voluntarily. That results in an increase in arterial partial oxygen pressure (PO2), which
produces an urge to breathe. When eventually PCO2 reaches the high enough level it overrides the
conscious influences from the cortex and stimulates the inspiratory system. If one holds his breath long
enough to decrease PO2 to a very low level one may loose his consciousness. In an unconscious person
automatic control of the respiration takes over and the normal breathing resumes.
Other parts of the brain (limbic system, hypothalamus) can also alter the breathing pattern e.g.
affective states, strong emotions such as rage and fear. In addition, stimulation of touch, thermal and
pain receptors can also stimulate the respiratory system.
9
RESPIRATORY MUSCLES: Diaphragm, intercostal muscles and the other accessory respiratory
muscles work in co-ordination for normal breathing under central controller. There is evidence
suggesting that in premature new-born babies this co-ordination is not mature enough and this could be
responsible for the sudden infant death syndrome.
RESPIRATORY MUSCLES: Diaphragm, intercostal muscles and the other accessory respiratory
muscles work in co-ordination for normal breathing under central controller. There is evidence
suggesting that in premature new-born babies this co-ordination is not mature enough and this could be
responsible for the sudden infant death syndrome.

10
SENSORS:
1.MECHANORECEPTORS: These receptors are placed in the walls of bronchi and bronchioles of
the lung and the main function of these receptors is to prevent the overinflation of the lungs. Inflation
of the lungs activates these receptors and activation of the stretch receptors in turn inhibits the
neurones in inspiratory centre via vagus nerve. When the expiration starts activation of the stretch
receptors gradually ceases allowing neurones in the inspiratory neurones become active again. This
phenomenon is called Hering-Breuer Reflex. It is particularly important for infants. In adults it is
functional only during exercise when the tidal volume is larger than normal.
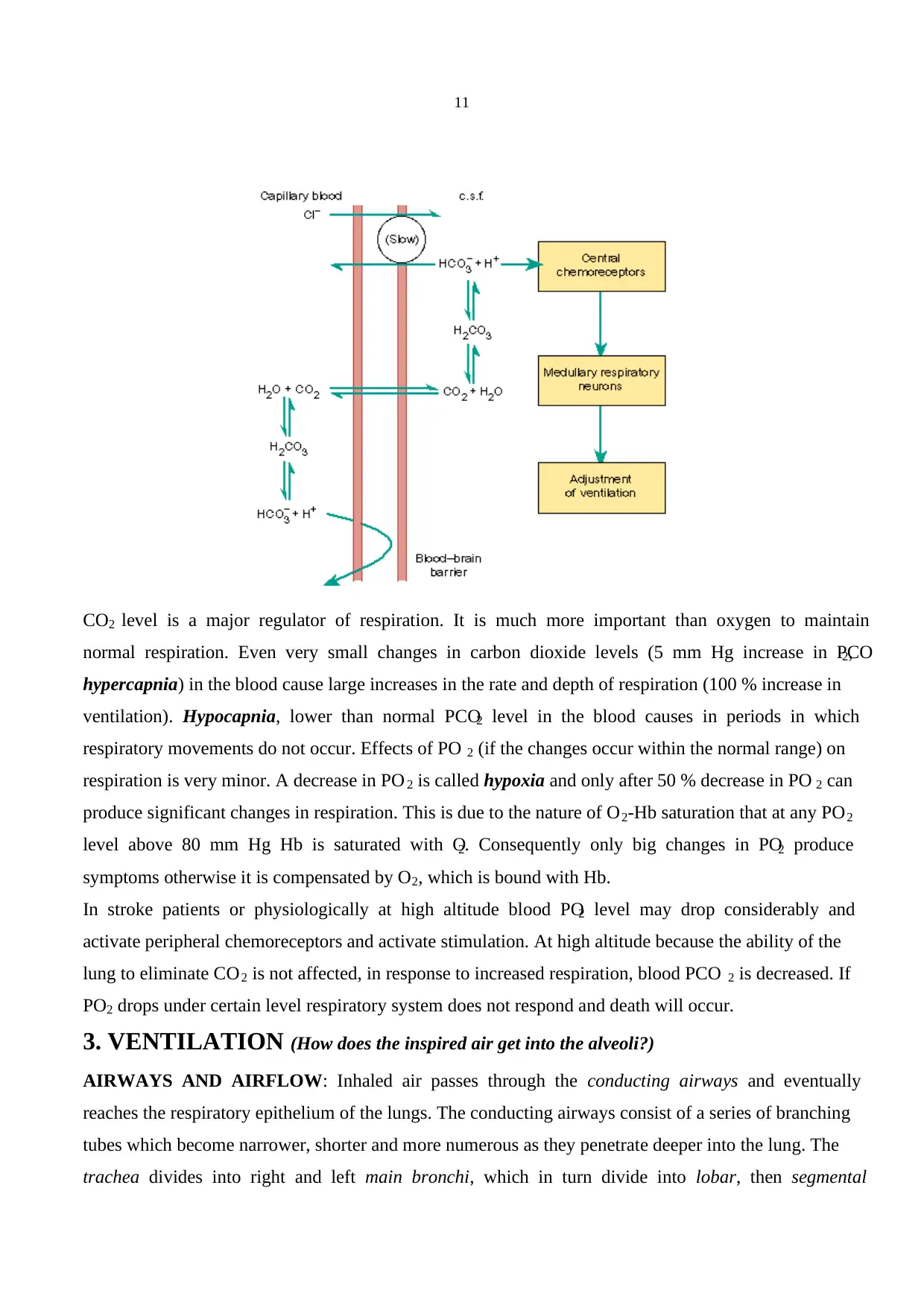
2.CHEMORECEPTORS: The respiratory system maintains concentrations of O 2, CO2 and the pH of
the body fluids within the normal range of values. Any deviation from these values has a marked
influence on the respiration. Chemoreceptors are specialised neurones activated by changes in O 2 or
CO2 levels in the blood and the brain tissue, respectively. They are involved in the regulation of
respiration according to the changes in PO2 and pH. O2-sensitive chemoreceptors (Peripheral
chemoreceptors) are located at the bifurcation of the carotid artery in the neck and the aortic arch.
They are small vascular sensory organs encapsulated with the connective tissue. They are connected to
the respiratory centre in the medulla by glossopharingeal nerve (carotid body chemoreceptors) and the
vagus nerve (aortic body). Central chemoreceptors are located bilaterally in the chemosensitive area of
the medulla oblongata and exposed to the cerebrospinal fluid (CSF), local blood flow and local
metabolism. They actually respond to changes in H + concentration in these compartments. When the
blood partial PCO2 is increased CO2 diffuses into the CSF from cerebral vessels and liberates H+.
(When CO2 combines with water forms carbonic acid and liberates H+ and HCO3-).
CO2 + H2O ↔ H2CO3
H2CO3 ↔ HCO -3 + H+
An increase in H+ stimulates chemoreceptors resulting in hyperventilation which in turn reduces PCO 2
in the blood and therefore in the CSF. Cerebral vasodilatation always accompanies an increased PCO 2
and enhances the diffusion of CO2 into the CSF. Because CSF has less protein than blood it has a
much lower buffering capacity. As a result changes in pH for a given change in PCO 2 is always bigger
than the change in blood.
SENSORS:
1.MECHANORECEPTORS: These receptors are placed in the walls of bronchi and bronchioles of
the lung and the main function of these receptors is to prevent the overinflation of the lungs. Inflation
of the lungs activates these receptors and activation of the stretch receptors in turn inhibits the
neurones in inspiratory centre via vagus nerve. When the expiration starts activation of the stretch
receptors gradually ceases allowing neurones in the inspiratory neurones become active again. This
phenomenon is called Hering-Breuer Reflex. It is particularly important for infants. In adults it is
functional only during exercise when the tidal volume is larger than normal.
2.CHEMORECEPTORS: The respiratory system maintains concentrations of O 2, CO2 and the pH of
the body fluids within the normal range of values. Any deviation from these values has a marked
influence on the respiration. Chemoreceptors are specialised neurones activated by changes in O 2 or
CO2 levels in the blood and the brain tissue, respectively. They are involved in the regulation of
respiration according to the changes in PO2 and pH. O2-sensitive chemoreceptors (Peripheral
chemoreceptors) are located at the bifurcation of the carotid artery in the neck and the aortic arch.
They are small vascular sensory organs encapsulated with the connective tissue. They are connected to
the respiratory centre in the medulla by glossopharingeal nerve (carotid body chemoreceptors) and the
vagus nerve (aortic body). Central chemoreceptors are located bilaterally in the chemosensitive area of
the medulla oblongata and exposed to the cerebrospinal fluid (CSF), local blood flow and local
metabolism. They actually respond to changes in H + concentration in these compartments. When the
blood partial PCO2 is increased CO2 diffuses into the CSF from cerebral vessels and liberates H+.
(When CO2 combines with water forms carbonic acid and liberates H+ and HCO3-).
CO2 + H2O ↔ H2CO3
H2CO3 ↔ HCO -3 + H+
An increase in H+ stimulates chemoreceptors resulting in hyperventilation which in turn reduces PCO 2
in the blood and therefore in the CSF. Cerebral vasodilatation always accompanies an increased PCO 2
and enhances the diffusion of CO2 into the CSF. Because CSF has less protein than blood it has a
much lower buffering capacity. As a result changes in pH for a given change in PCO 2 is always bigger
than the change in blood.
Secure Best Marks with AI Grader
Need help grading? Try our AI Grader for instant feedback on your assignments.
11
CO2 level is a major regulator of respiration. It is much more important than oxygen to maintain
normal respiration. Even very small changes in carbon dioxide levels (5 mm Hg increase in PCO2,
hypercapnia) in the blood cause large increases in the rate and depth of respiration (100 % increase in
ventilation). Hypocapnia, lower than normal PCO2 level in the blood causes in periods in which
respiratory movements do not occur. Effects of PO 2 (if the changes occur within the normal range) on
respiration is very minor. A decrease in PO 2 is called hypoxia and only after 50 % decrease in PO 2 can
produce significant changes in respiration. This is due to the nature of O2-Hb saturation that at any PO2
level above 80 mm Hg Hb is saturated with O2. Consequently only big changes in PO2 produce
symptoms otherwise it is compensated by O2, which is bound with Hb.
In stroke patients or physiologically at high altitude blood PO2 level may drop considerably and
activate peripheral chemoreceptors and activate stimulation. At high altitude because the ability of the
lung to eliminate CO2 is not affected, in response to increased respiration, blood PCO 2 is decreased. If
PO2 drops under certain level respiratory system does not respond and death will occur.
3. VENTILATION (How does the inspired air get into the alveoli?)
AIRWAYS AND AIRFLOW: Inhaled air passes through the conducting airways and eventually
reaches the respiratory epithelium of the lungs. The conducting airways consist of a series of branching
tubes which become narrower, shorter and more numerous as they penetrate deeper into the lung. The
trachea divides into right and left main bronchi, which in turn divide into lobar, then segmental
CO2 level is a major regulator of respiration. It is much more important than oxygen to maintain
normal respiration. Even very small changes in carbon dioxide levels (5 mm Hg increase in PCO2,
hypercapnia) in the blood cause large increases in the rate and depth of respiration (100 % increase in
ventilation). Hypocapnia, lower than normal PCO2 level in the blood causes in periods in which
respiratory movements do not occur. Effects of PO 2 (if the changes occur within the normal range) on
respiration is very minor. A decrease in PO 2 is called hypoxia and only after 50 % decrease in PO 2 can
produce significant changes in respiration. This is due to the nature of O2-Hb saturation that at any PO2
level above 80 mm Hg Hb is saturated with O2. Consequently only big changes in PO2 produce
symptoms otherwise it is compensated by O2, which is bound with Hb.
In stroke patients or physiologically at high altitude blood PO2 level may drop considerably and
activate peripheral chemoreceptors and activate stimulation. At high altitude because the ability of the
lung to eliminate CO2 is not affected, in response to increased respiration, blood PCO 2 is decreased. If
PO2 drops under certain level respiratory system does not respond and death will occur.
3. VENTILATION (How does the inspired air get into the alveoli?)
AIRWAYS AND AIRFLOW: Inhaled air passes through the conducting airways and eventually
reaches the respiratory epithelium of the lungs. The conducting airways consist of a series of branching
tubes which become narrower, shorter and more numerous as they penetrate deeper into the lung. The
trachea divides into right and left main bronchi, which in turn divide into lobar, then segmental
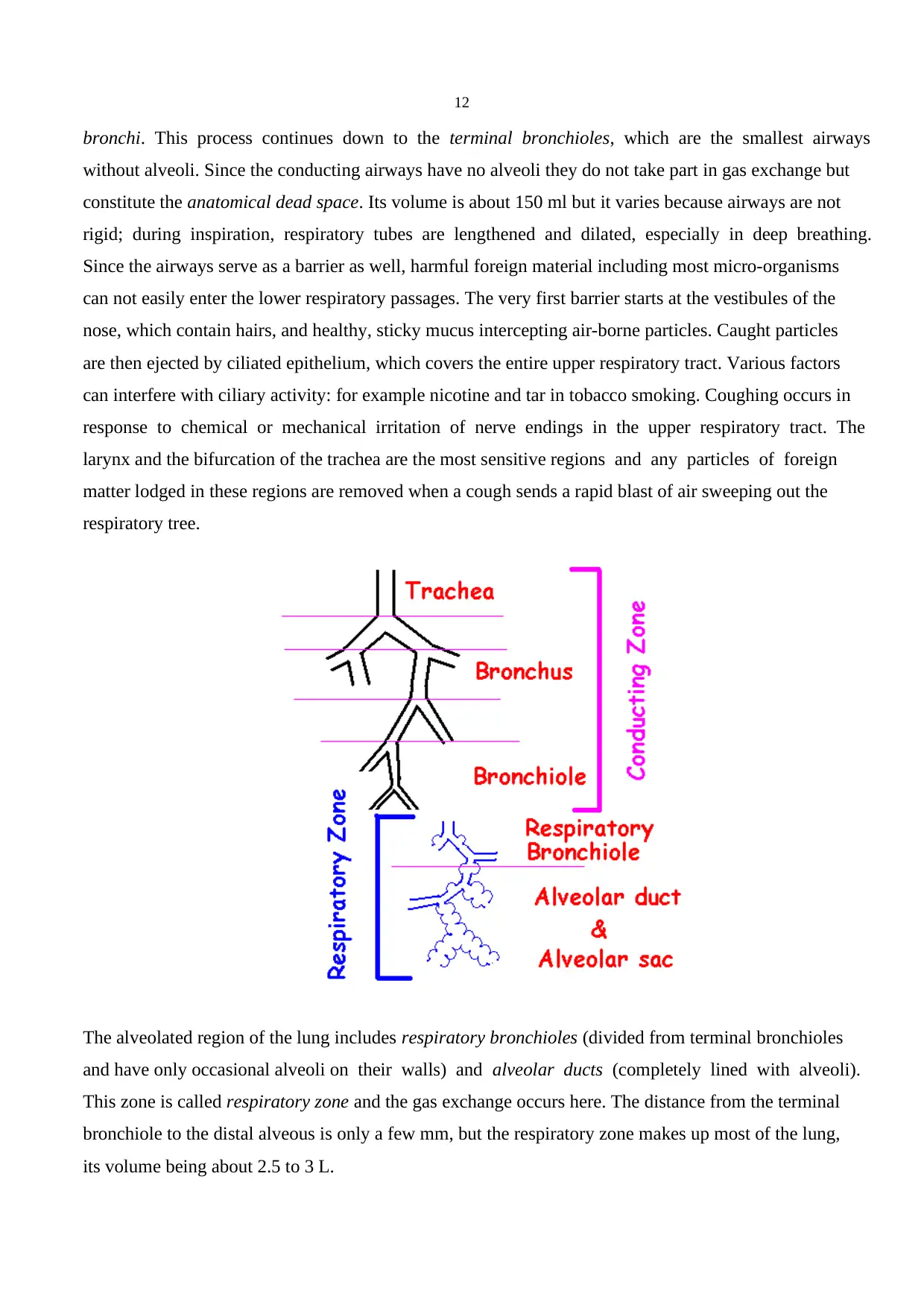
12
bronchi. This process continues down to the terminal bronchioles, which are the smallest airways
without alveoli. Since the conducting airways have no alveoli they do not take part in gas exchange but
constitute the anatomical dead space. Its volume is about 150 ml but it varies because airways are not
rigid; during inspiration, respiratory tubes are lengthened and dilated, especially in deep breathing.
Since the airways serve as a barrier as well, harmful foreign material including most micro-organisms
can not easily enter the lower respiratory passages. The very first barrier starts at the vestibules of the
nose, which contain hairs, and healthy, sticky mucus intercepting air-borne particles. Caught particles
are then ejected by ciliated epithelium, which covers the entire upper respiratory tract. Various factors
can interfere with ciliary activity: for example nicotine and tar in tobacco smoking. Coughing occurs in
response to chemical or mechanical irritation of nerve endings in the upper respiratory tract. The
larynx and the bifurcation of the trachea are the most sensitive regions and any particles of foreign
matter lodged in these regions are removed when a cough sends a rapid blast of air sweeping out the
respiratory tree.
The alveolated region of the lung includes respiratory bronchioles (divided from terminal bronchioles
and have only occasional alveoli on their walls) and alveolar ducts (completely lined with alveoli).
This zone is called respiratory zone and the gas exchange occurs here. The distance from the terminal
bronchiole to the distal alveous is only a few mm, but the respiratory zone makes up most of the lung,
its volume being about 2.5 to 3 L.
bronchi. This process continues down to the terminal bronchioles, which are the smallest airways
without alveoli. Since the conducting airways have no alveoli they do not take part in gas exchange but
constitute the anatomical dead space. Its volume is about 150 ml but it varies because airways are not
rigid; during inspiration, respiratory tubes are lengthened and dilated, especially in deep breathing.
Since the airways serve as a barrier as well, harmful foreign material including most micro-organisms
can not easily enter the lower respiratory passages. The very first barrier starts at the vestibules of the
nose, which contain hairs, and healthy, sticky mucus intercepting air-borne particles. Caught particles
are then ejected by ciliated epithelium, which covers the entire upper respiratory tract. Various factors
can interfere with ciliary activity: for example nicotine and tar in tobacco smoking. Coughing occurs in
response to chemical or mechanical irritation of nerve endings in the upper respiratory tract. The
larynx and the bifurcation of the trachea are the most sensitive regions and any particles of foreign
matter lodged in these regions are removed when a cough sends a rapid blast of air sweeping out the
respiratory tree.
The alveolated region of the lung includes respiratory bronchioles (divided from terminal bronchioles
and have only occasional alveoli on their walls) and alveolar ducts (completely lined with alveoli).
This zone is called respiratory zone and the gas exchange occurs here. The distance from the terminal
bronchiole to the distal alveous is only a few mm, but the respiratory zone makes up most of the lung,
its volume being about 2.5 to 3 L.

13
Blood is brought to the other side of the blood-gas barrier from the right heart by pulmonary arteries,
which also form a series of branching tubes leading to the pulmonary capillaries and back to the
pulmonary veins. The capillaries lie in the walls of the alveoli and form a dense network that the blood
continuously runs in the alveolar wall. At resting not all the capillaries are open but when the pressure
rises (e.g. exercise) recruitment of the close capillaries occurs. The diameter of a capillary segment is
about 10 μm, just large enough for a red blood cell. The pulmonary artery receives the whole output of
the right heart, but resistance of pulmonary circuit is very low. This enables the high blood flow to the
circuit.
4. LUNG VOLUMES AND PULMONARY FUNCTION TESTS:
Pulmonary function can be examined by the spirometry technique. Spirometers are the traditional
tools of the respiratory physiologists. The subject breathes into a closed system in which air is trapped
(bell). As the subject breathes air movement into or out of the mouthpiece causes the bell to rise
(inspiration) or fall (expiration). Corresponding movements of an attached pen register the change in
volume on a rotating drum recorder. From such a recording we could measure
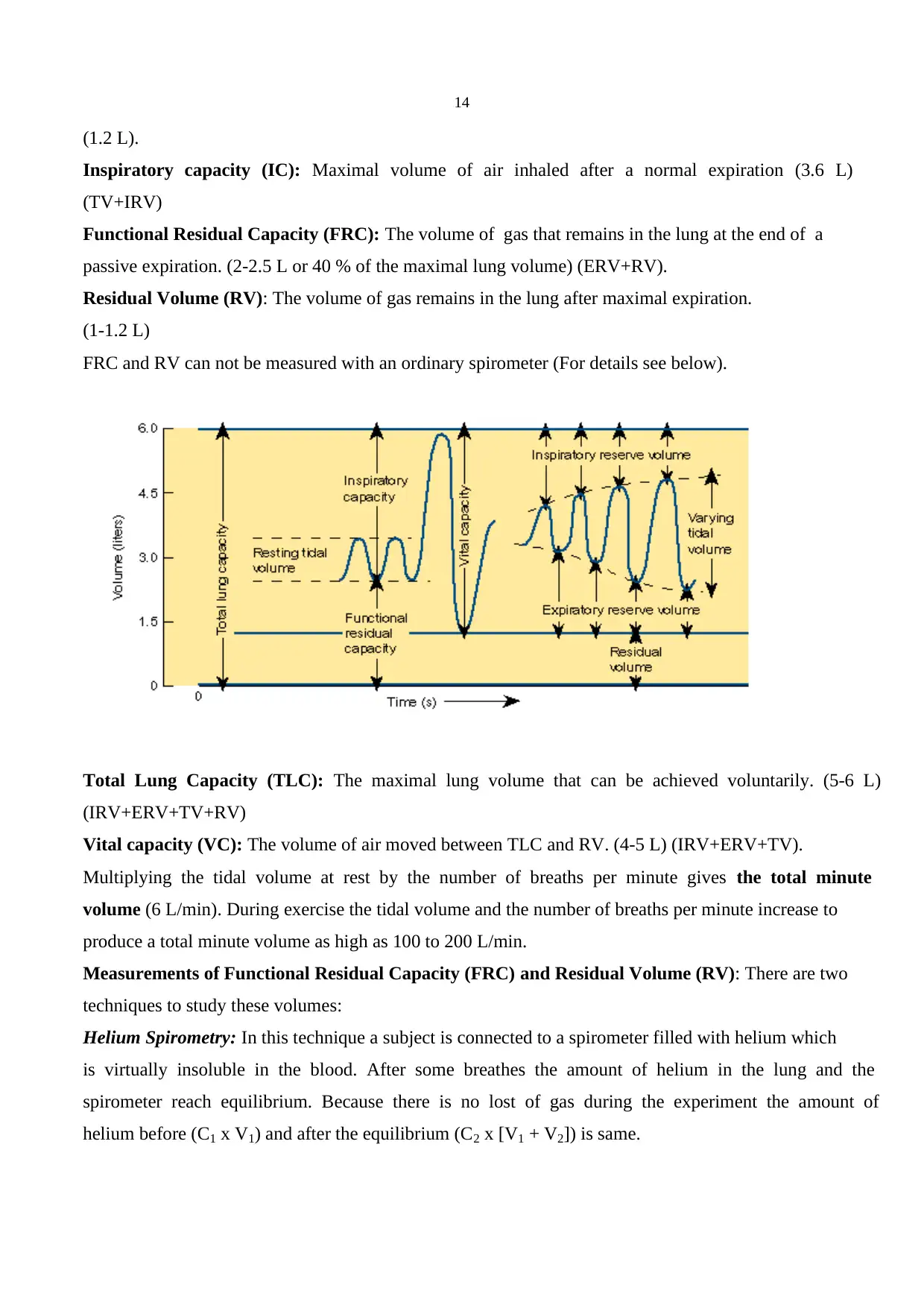
Tidal volume (TV): Volume of air inhaled or exhaled with each breath during normal breathing
(0.5 L).
Inspiratory reserve volume (IRV): Maximal volume of air inhaled at the end of a normal inspiration
(3 L)
Expiratory reserve volume (ERV): Maximal volume of air exhaled at the end of a tidal volume
Blood is brought to the other side of the blood-gas barrier from the right heart by pulmonary arteries,
which also form a series of branching tubes leading to the pulmonary capillaries and back to the
pulmonary veins. The capillaries lie in the walls of the alveoli and form a dense network that the blood
continuously runs in the alveolar wall. At resting not all the capillaries are open but when the pressure
rises (e.g. exercise) recruitment of the close capillaries occurs. The diameter of a capillary segment is
about 10 μm, just large enough for a red blood cell. The pulmonary artery receives the whole output of
the right heart, but resistance of pulmonary circuit is very low. This enables the high blood flow to the
circuit.
4. LUNG VOLUMES AND PULMONARY FUNCTION TESTS:
Pulmonary function can be examined by the spirometry technique. Spirometers are the traditional
tools of the respiratory physiologists. The subject breathes into a closed system in which air is trapped
(bell). As the subject breathes air movement into or out of the mouthpiece causes the bell to rise
(inspiration) or fall (expiration). Corresponding movements of an attached pen register the change in
volume on a rotating drum recorder. From such a recording we could measure
Tidal volume (TV): Volume of air inhaled or exhaled with each breath during normal breathing
(0.5 L).
Inspiratory reserve volume (IRV): Maximal volume of air inhaled at the end of a normal inspiration
(3 L)
Expiratory reserve volume (ERV): Maximal volume of air exhaled at the end of a tidal volume
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser
14
(1.2 L).
Inspiratory capacity (IC): Maximal volume of air inhaled after a normal expiration (3.6 L)
(TV+IRV)
Functional Residual Capacity (FRC): The volume of gas that remains in the lung at the end of a
passive expiration. (2-2.5 L or 40 % of the maximal lung volume) (ERV+RV).
Residual Volume (RV): The volume of gas remains in the lung after maximal expiration.
(1-1.2 L)
FRC and RV can not be measured with an ordinary spirometer (For details see below).
Total Lung Capacity (TLC): The maximal lung volume that can be achieved voluntarily. (5-6 L)
(IRV+ERV+TV+RV)
Vital capacity (VC): The volume of air moved between TLC and RV. (4-5 L) (IRV+ERV+TV).
Multiplying the tidal volume at rest by the number of breaths per minute gives the total minute
volume (6 L/min). During exercise the tidal volume and the number of breaths per minute increase to
produce a total minute volume as high as 100 to 200 L/min.
Measurements of Functional Residual Capacity (FRC) and Residual Volume (RV): There are two
techniques to study these volumes:
Helium Spirometry: In this technique a subject is connected to a spirometer filled with helium which
is virtually insoluble in the blood. After some breathes the amount of helium in the lung and the
spirometer reach equilibrium. Because there is no lost of gas during the experiment the amount of
helium before (C1 x V1) and after the equilibrium (C2 x [V1 + V2]) is same.
(1.2 L).
Inspiratory capacity (IC): Maximal volume of air inhaled after a normal expiration (3.6 L)
(TV+IRV)
Functional Residual Capacity (FRC): The volume of gas that remains in the lung at the end of a
passive expiration. (2-2.5 L or 40 % of the maximal lung volume) (ERV+RV).
Residual Volume (RV): The volume of gas remains in the lung after maximal expiration.
(1-1.2 L)
FRC and RV can not be measured with an ordinary spirometer (For details see below).
Total Lung Capacity (TLC): The maximal lung volume that can be achieved voluntarily. (5-6 L)
(IRV+ERV+TV+RV)
Vital capacity (VC): The volume of air moved between TLC and RV. (4-5 L) (IRV+ERV+TV).
Multiplying the tidal volume at rest by the number of breaths per minute gives the total minute
volume (6 L/min). During exercise the tidal volume and the number of breaths per minute increase to
produce a total minute volume as high as 100 to 200 L/min.
Measurements of Functional Residual Capacity (FRC) and Residual Volume (RV): There are two
techniques to study these volumes:
Helium Spirometry: In this technique a subject is connected to a spirometer filled with helium which
is virtually insoluble in the blood. After some breathes the amount of helium in the lung and the
spirometer reach equilibrium. Because there is no lost of gas during the experiment the amount of
helium before (C1 x V1) and after the equilibrium (C2 x [V1 + V2]) is same.

15
V1 = C2 x (V1 + V2)
V2 = V1 x (C1 – C2) / C2
V2 = FRC
Another way of measuring FRC is with a body plethysmograph. It is a big airtight box in which the
subject sits. At the end of a normal expiration, the mouthpiece is shut and the subject makes
respiratory efforts.
When the subject makes an inspiratory effort against a closed airway s/he slightly increases the volume
of his/her lung, airway pressure decreases and the box pressure increases:
P1xV1 = P2x(V1-∆V)
The pressure in the box before (P1) and after (P2) the respiratory efforts,
V: Volume in the box before the respiratory efforts and ∆V can be measured.
The Boyle’s law can also be applied to the gas in the lung:
P3 x V2 = P4 x (V2 + ∆ V)
V2 = FRC
P3,4: Mouth pressures before (P3) and after (P4) the respiratory efforts.
If the measurement is done following a forced expiration
V2 = RV
V1 = C2 x (V1 + V2)
V2 = V1 x (C1 – C2) / C2
V2 = FRC
Another way of measuring FRC is with a body plethysmograph. It is a big airtight box in which the
subject sits. At the end of a normal expiration, the mouthpiece is shut and the subject makes
respiratory efforts.
When the subject makes an inspiratory effort against a closed airway s/he slightly increases the volume
of his/her lung, airway pressure decreases and the box pressure increases:
P1xV1 = P2x(V1-∆V)
The pressure in the box before (P1) and after (P2) the respiratory efforts,
V: Volume in the box before the respiratory efforts and ∆V can be measured.
The Boyle’s law can also be applied to the gas in the lung:
P3 x V2 = P4 x (V2 + ∆ V)
V2 = FRC
P3,4: Mouth pressures before (P3) and after (P4) the respiratory efforts.
If the measurement is done following a forced expiration
V2 = RV

16
In contrast to the helium technique, which measures only the ventilated air, the body plethysmograph
measures the total volume in the gas in the volume including the gas trapped in the airways (if there is
any). Normally measurements with these techniques are similar. However, the difference is increased
in the presence of lung diseases.
TOTAL VENTILATION: The total volume of the gas leaving the lung per unit time. If TV is 500 ml
and there are approximately 15 breaths/min the total volume of the gas leaving the lung, total
ventilation will be 500 x 15 = 7500 ml/min. It can be measured by having the subject breath through a
valve that separates the inspired air from expired air and collecting the expired air.
ALVEOLAR VENTILATION: The volume of the gas reaching the respiratory zone of the airways.
However, not all of the total ventilation volume reaches the alveoli. 150 ml of the TV (500 ml) is left
behind in the airways, which does not contain alveoli, therefore does not contribute the diffusion
(Anatomic death space). Thus, the volume of gas entering the respiratory zone, alveolar ventilation, is
(500-150) x 15 = 5250 ml/min. The measurement of alveolar ventilation is more difficult. One way is
to measure the volume of anatomic dead space and calculate the dead space ventilation. This then
subtracted from the total ventilation.
Alveolar ventilation = Total ventilation – Anatomic death space ventilation
Anatomic dead space ventilation = Anatomic dead space volume x respiration frequency
VE: total expiration volume
VT: Tidal volume
VD: Dead Space volume
VA: volume of alveolar gas during tidal breathing
V: volume per unit time
VT = VD + VA
In contrast to the helium technique, which measures only the ventilated air, the body plethysmograph
measures the total volume in the gas in the volume including the gas trapped in the airways (if there is
any). Normally measurements with these techniques are similar. However, the difference is increased
in the presence of lung diseases.
TOTAL VENTILATION: The total volume of the gas leaving the lung per unit time. If TV is 500 ml
and there are approximately 15 breaths/min the total volume of the gas leaving the lung, total
ventilation will be 500 x 15 = 7500 ml/min. It can be measured by having the subject breath through a
valve that separates the inspired air from expired air and collecting the expired air.
ALVEOLAR VENTILATION: The volume of the gas reaching the respiratory zone of the airways.
However, not all of the total ventilation volume reaches the alveoli. 150 ml of the TV (500 ml) is left
behind in the airways, which does not contain alveoli, therefore does not contribute the diffusion
(Anatomic death space). Thus, the volume of gas entering the respiratory zone, alveolar ventilation, is
(500-150) x 15 = 5250 ml/min. The measurement of alveolar ventilation is more difficult. One way is
to measure the volume of anatomic dead space and calculate the dead space ventilation. This then
subtracted from the total ventilation.
Alveolar ventilation = Total ventilation – Anatomic death space ventilation
Anatomic dead space ventilation = Anatomic dead space volume x respiration frequency
VE: total expiration volume
VT: Tidal volume
VD: Dead Space volume
VA: volume of alveolar gas during tidal breathing
V: volume per unit time
VT = VD + VA
Secure Best Marks with AI Grader
Need help grading? Try our AI Grader for instant feedback on your assignments.

17
(VT x n) = (VD x n) + (VA x n)
V: volume per unit time
VE: Expired total ventilation
VD: dead space ventilation
VA: alveolar ventilation
VE = VD + VA
VA = VE - VD
The disadvantage of this method is it is not very easy to determine dead space volume without a
considerable error.
Another way of measuring the alveolar ventilation is from concentration of CO2 in expired air. Since
the amount of CO2 in the inspired air is negligible and no gas exchange occurs, we could assume that
there is CO2 in the anatomic dead space. Therefor CO2 in the expired air comes from alveoli.
VCO2 = VA x % CO2 / 100
VCO2 = the volume of CO2 exhaled per unit time.
VA = ( VCO2 x 100 ) / % CO2
% CO2 = Fractional CO2 concentration = FCO2
ANATOMICAL DEAD SPACE: Volume of the conducting airways. It is approximately 150 ml but
its volume increases with large inspiration and depends on the size and the posture of the subject.
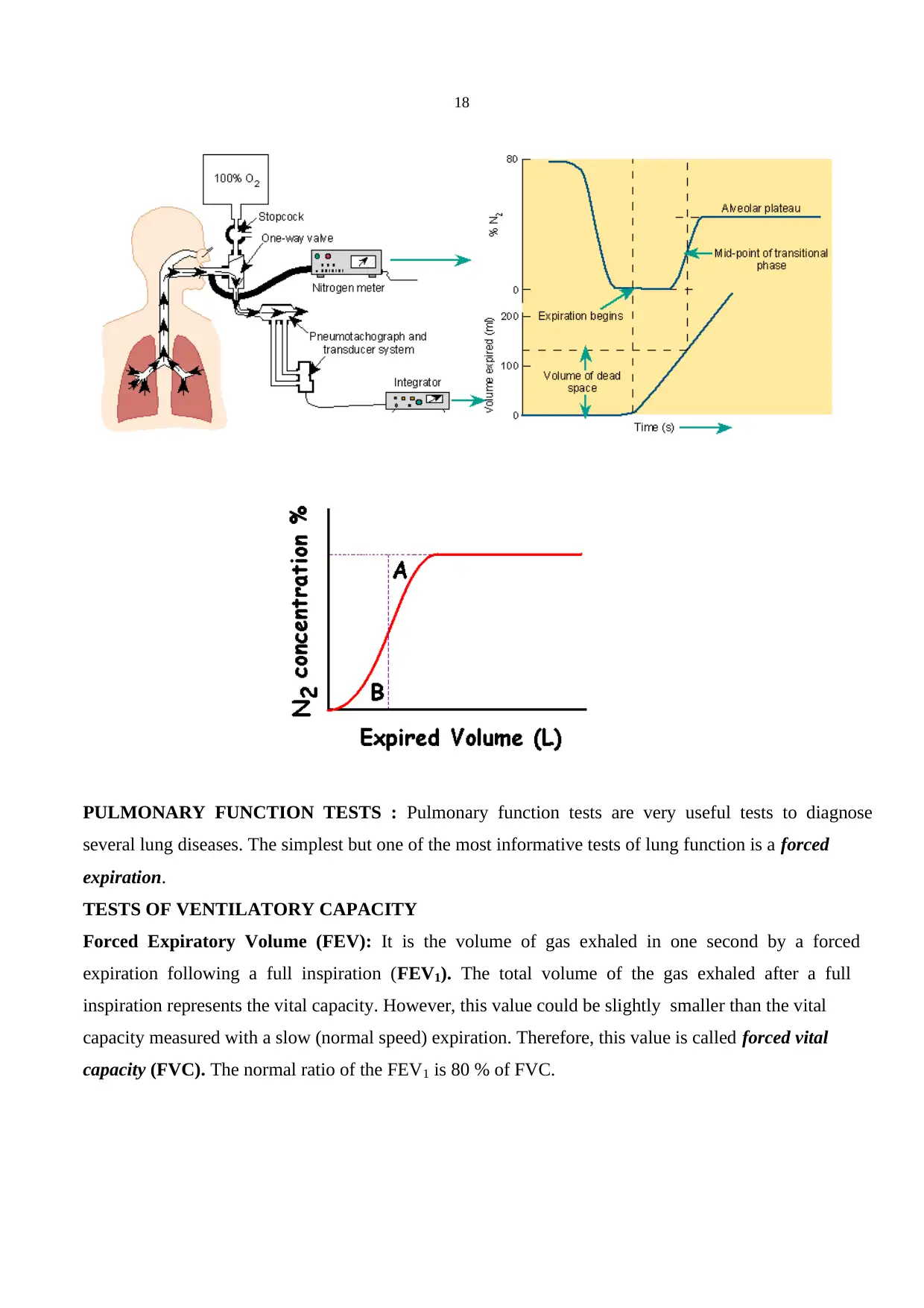
Measurement of dead space: Fowler’s method: The subject breaths pure oxygen through a valve box
and a rapid nitrogen analyser samples and measures the nitrogen concentration in the expired air. After
a single inspiration of pure oxygen (100 %) nitrogen concentration in the expired air is increased as the
gas in the dead space is washed by pure oxygen. Nitrogen concentration quickly reaches a plateau level
(alveolar plateau). The dead space is found by plotting nitrogen concentration against the expired
volume. The expired volume up to the vertical line drawn such that area A = area B represents the
anatomical dead space volume.
(VT x n) = (VD x n) + (VA x n)
V: volume per unit time
VE: Expired total ventilation
VD: dead space ventilation
VA: alveolar ventilation
VE = VD + VA
VA = VE - VD
The disadvantage of this method is it is not very easy to determine dead space volume without a
considerable error.
Another way of measuring the alveolar ventilation is from concentration of CO2 in expired air. Since
the amount of CO2 in the inspired air is negligible and no gas exchange occurs, we could assume that
there is CO2 in the anatomic dead space. Therefor CO2 in the expired air comes from alveoli.
VCO2 = VA x % CO2 / 100
VCO2 = the volume of CO2 exhaled per unit time.
VA = ( VCO2 x 100 ) / % CO2
% CO2 = Fractional CO2 concentration = FCO2
ANATOMICAL DEAD SPACE: Volume of the conducting airways. It is approximately 150 ml but
its volume increases with large inspiration and depends on the size and the posture of the subject.
Measurement of dead space: Fowler’s method: The subject breaths pure oxygen through a valve box
and a rapid nitrogen analyser samples and measures the nitrogen concentration in the expired air. After
a single inspiration of pure oxygen (100 %) nitrogen concentration in the expired air is increased as the
gas in the dead space is washed by pure oxygen. Nitrogen concentration quickly reaches a plateau level
(alveolar plateau). The dead space is found by plotting nitrogen concentration against the expired
volume. The expired volume up to the vertical line drawn such that area A = area B represents the
anatomical dead space volume.
18
PULMONARY FUNCTION TESTS : Pulmonary function tests are very useful tests to diagnose
several lung diseases. The simplest but one of the most informative tests of lung function is a forced
expiration.
TESTS OF VENTILATORY CAPACITY
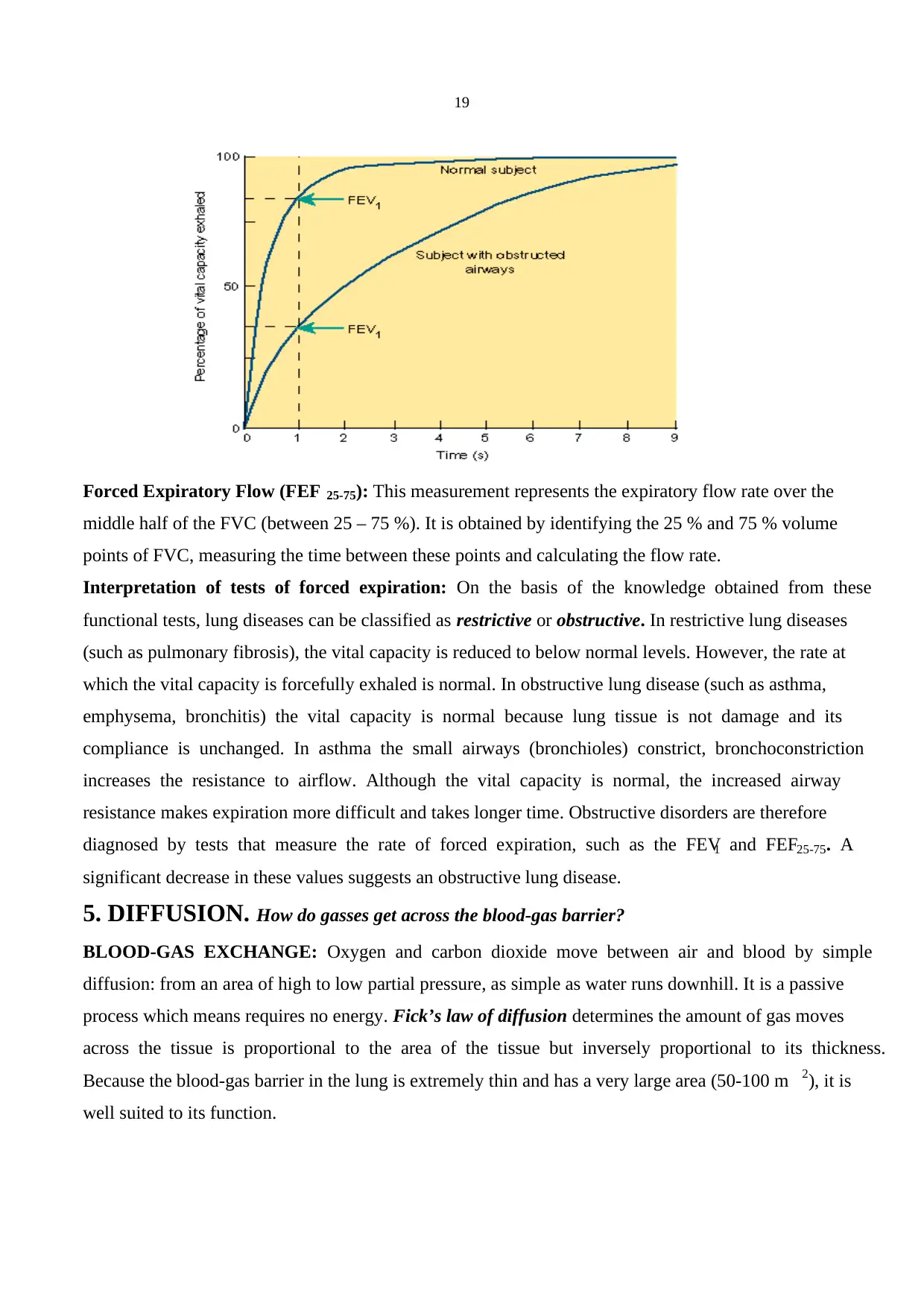
Forced Expiratory Volume (FEV): It is the volume of gas exhaled in one second by a forced
expiration following a full inspiration (FEV1). The total volume of the gas exhaled after a full
inspiration represents the vital capacity. However, this value could be slightly smaller than the vital
capacity measured with a slow (normal speed) expiration. Therefore, this value is called forced vital
capacity (FVC). The normal ratio of the FEV1 is 80 % of FVC.
PULMONARY FUNCTION TESTS : Pulmonary function tests are very useful tests to diagnose
several lung diseases. The simplest but one of the most informative tests of lung function is a forced
expiration.
TESTS OF VENTILATORY CAPACITY
Forced Expiratory Volume (FEV): It is the volume of gas exhaled in one second by a forced
expiration following a full inspiration (FEV1). The total volume of the gas exhaled after a full
inspiration represents the vital capacity. However, this value could be slightly smaller than the vital
capacity measured with a slow (normal speed) expiration. Therefore, this value is called forced vital
capacity (FVC). The normal ratio of the FEV1 is 80 % of FVC.
19
Forced Expiratory Flow (FEF 25-75): This measurement represents the expiratory flow rate over the
middle half of the FVC (between 25 – 75 %). It is obtained by identifying the 25 % and 75 % volume
points of FVC, measuring the time between these points and calculating the flow rate.
Interpretation of tests of forced expiration: On the basis of the knowledge obtained from these
functional tests, lung diseases can be classified as restrictive or obstructive. In restrictive lung diseases
(such as pulmonary fibrosis), the vital capacity is reduced to below normal levels. However, the rate at
which the vital capacity is forcefully exhaled is normal. In obstructive lung disease (such as asthma,
emphysema, bronchitis) the vital capacity is normal because lung tissue is not damage and its
compliance is unchanged. In asthma the small airways (bronchioles) constrict, bronchoconstriction
increases the resistance to airflow. Although the vital capacity is normal, the increased airway
resistance makes expiration more difficult and takes longer time. Obstructive disorders are therefore
diagnosed by tests that measure the rate of forced expiration, such as the FEV1 and FEF25-75. A
significant decrease in these values suggests an obstructive lung disease.
5. DIFFUSION. How do gasses get across the blood-gas barrier?
BLOOD-GAS EXCHANGE: Oxygen and carbon dioxide move between air and blood by simple
diffusion: from an area of high to low partial pressure, as simple as water runs downhill. It is a passive
process which means requires no energy. Fick’s law of diffusion determines the amount of gas moves
across the tissue is proportional to the area of the tissue but inversely proportional to its thickness.
Because the blood-gas barrier in the lung is extremely thin and has a very large area (50-100 m 2), it is
well suited to its function.
Forced Expiratory Flow (FEF 25-75): This measurement represents the expiratory flow rate over the
middle half of the FVC (between 25 – 75 %). It is obtained by identifying the 25 % and 75 % volume
points of FVC, measuring the time between these points and calculating the flow rate.
Interpretation of tests of forced expiration: On the basis of the knowledge obtained from these
functional tests, lung diseases can be classified as restrictive or obstructive. In restrictive lung diseases
(such as pulmonary fibrosis), the vital capacity is reduced to below normal levels. However, the rate at
which the vital capacity is forcefully exhaled is normal. In obstructive lung disease (such as asthma,
emphysema, bronchitis) the vital capacity is normal because lung tissue is not damage and its
compliance is unchanged. In asthma the small airways (bronchioles) constrict, bronchoconstriction
increases the resistance to airflow. Although the vital capacity is normal, the increased airway
resistance makes expiration more difficult and takes longer time. Obstructive disorders are therefore
diagnosed by tests that measure the rate of forced expiration, such as the FEV1 and FEF25-75. A
significant decrease in these values suggests an obstructive lung disease.
5. DIFFUSION. How do gasses get across the blood-gas barrier?
BLOOD-GAS EXCHANGE: Oxygen and carbon dioxide move between air and blood by simple
diffusion: from an area of high to low partial pressure, as simple as water runs downhill. It is a passive
process which means requires no energy. Fick’s law of diffusion determines the amount of gas moves
across the tissue is proportional to the area of the tissue but inversely proportional to its thickness.
Because the blood-gas barrier in the lung is extremely thin and has a very large area (50-100 m 2), it is
well suited to its function.
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser
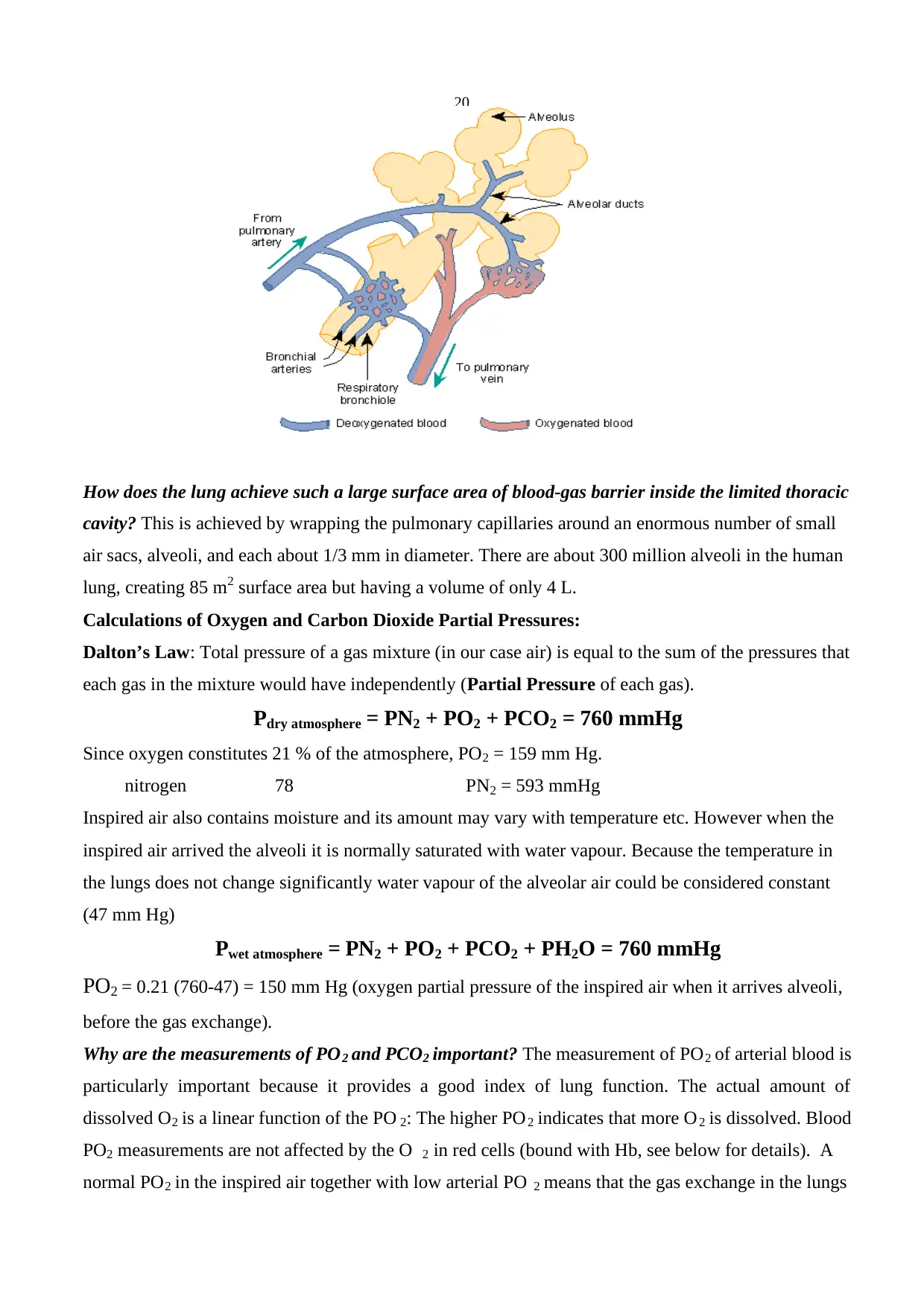
20
How does the lung achieve such a large surface area of blood-gas barrier inside the limited thoracic
cavity? This is achieved by wrapping the pulmonary capillaries around an enormous number of small
air sacs, alveoli, and each about 1/3 mm in diameter. There are about 300 million alveoli in the human
lung, creating 85 m2 surface area but having a volume of only 4 L.
Calculations of Oxygen and Carbon Dioxide Partial Pressures:
Dalton’s Law: Total pressure of a gas mixture (in our case air) is equal to the sum of the pressures that
each gas in the mixture would have independently (Partial Pressure of each gas).
Pdry atmosphere = PN2 + PO2 + PCO2 = 760 mmHg
Since oxygen constitutes 21 % of the atmosphere, PO2 = 159 mm Hg.
nitrogen 78 PN2 = 593 mmHg
Inspired air also contains moisture and its amount may vary with temperature etc. However when the
inspired air arrived the alveoli it is normally saturated with water vapour. Because the temperature in
the lungs does not change significantly water vapour of the alveolar air could be considered constant
(47 mm Hg)
Pwet atmosphere = PN2 + PO2 + PCO2 + PH2O = 760 mmHg
PO2 = 0.21 (760-47) = 150 mm Hg (oxygen partial pressure of the inspired air when it arrives alveoli,
before the gas exchange).
Why are the measurements of PO2 and PCO2 important? The measurement of PO2 of arterial blood is
particularly important because it provides a good index of lung function. The actual amount of
dissolved O2 is a linear function of the PO 2: The higher PO2 indicates that more O2 is dissolved. Blood
PO2 measurements are not affected by the O 2 in red cells (bound with Hb, see below for details). A
normal PO2 in the inspired air together with low arterial PO 2 means that the gas exchange in the lungs
How does the lung achieve such a large surface area of blood-gas barrier inside the limited thoracic
cavity? This is achieved by wrapping the pulmonary capillaries around an enormous number of small
air sacs, alveoli, and each about 1/3 mm in diameter. There are about 300 million alveoli in the human
lung, creating 85 m2 surface area but having a volume of only 4 L.
Calculations of Oxygen and Carbon Dioxide Partial Pressures:
Dalton’s Law: Total pressure of a gas mixture (in our case air) is equal to the sum of the pressures that
each gas in the mixture would have independently (Partial Pressure of each gas).
Pdry atmosphere = PN2 + PO2 + PCO2 = 760 mmHg
Since oxygen constitutes 21 % of the atmosphere, PO2 = 159 mm Hg.
nitrogen 78 PN2 = 593 mmHg
Inspired air also contains moisture and its amount may vary with temperature etc. However when the
inspired air arrived the alveoli it is normally saturated with water vapour. Because the temperature in
the lungs does not change significantly water vapour of the alveolar air could be considered constant
(47 mm Hg)
Pwet atmosphere = PN2 + PO2 + PCO2 + PH2O = 760 mmHg
PO2 = 0.21 (760-47) = 150 mm Hg (oxygen partial pressure of the inspired air when it arrives alveoli,
before the gas exchange).
Why are the measurements of PO2 and PCO2 important? The measurement of PO2 of arterial blood is
particularly important because it provides a good index of lung function. The actual amount of
dissolved O2 is a linear function of the PO 2: The higher PO2 indicates that more O2 is dissolved. Blood
PO2 measurements are not affected by the O 2 in red cells (bound with Hb, see below for details). A
normal PO2 in the inspired air together with low arterial PO 2 means that the gas exchange in the lungs

21
is impaired. In summary, the measurement of PO 2 is important for (1) treating patients with pulmonary
diseases (2) performing safe surgery (when anaesthesia is used) (3) the care of premature babies with
respiratory distress syndrome.
Inspired Air Alveolar Air
H2O Variable 47 mm Hg
CO2 0.3 mm Hg 40 mm Hg
O2 159 mm Hg 105 mm Hg
N2 601 mm Hg 568 mm Hg
Total Pressure 760 mm Hg 760 mm Hg
DIFFUSION AND PERFUSION LIMITATIONS: How does oxygen get into the circulation? To
answer this question we will first examine two extreme examples: If a subject breathes CO (carbon
monoxide) because CO moves rapidly across the blood-gas barrier, the content of CO in red blood
cells is increased. However, CO forms very tight bonds with Hb that even though large amount of CO
is taken up by the red blood cells almost no increase in the CO partial pressure is observed. Therefore,
the amount of CO that gets unto the blood is limited by the diffusion properties of the blood-gas barrier
and not by the amount of blood available: diffusion limited.
The other extreme example is nitrous oxide: Nitrous oxide diffuses across the barrier but forms no
combination with Hb. As a result its partial pressure rises very rapidly. The amount of nitrous oxide
taken up by blood depends on the amount of blood available: perfusion limited. The time course of O 2
transfer is in between. It does bind to Hb but nothing like the avidity of CO. In normal conditions
capillary PO2 reaches that of alveolar gas when the red cell is about 1/3 of the way along the capillary.
is impaired. In summary, the measurement of PO 2 is important for (1) treating patients with pulmonary
diseases (2) performing safe surgery (when anaesthesia is used) (3) the care of premature babies with
respiratory distress syndrome.
Inspired Air Alveolar Air
H2O Variable 47 mm Hg
CO2 0.3 mm Hg 40 mm Hg
O2 159 mm Hg 105 mm Hg
N2 601 mm Hg 568 mm Hg
Total Pressure 760 mm Hg 760 mm Hg
DIFFUSION AND PERFUSION LIMITATIONS: How does oxygen get into the circulation? To
answer this question we will first examine two extreme examples: If a subject breathes CO (carbon
monoxide) because CO moves rapidly across the blood-gas barrier, the content of CO in red blood
cells is increased. However, CO forms very tight bonds with Hb that even though large amount of CO
is taken up by the red blood cells almost no increase in the CO partial pressure is observed. Therefore,
the amount of CO that gets unto the blood is limited by the diffusion properties of the blood-gas barrier
and not by the amount of blood available: diffusion limited.
The other extreme example is nitrous oxide: Nitrous oxide diffuses across the barrier but forms no
combination with Hb. As a result its partial pressure rises very rapidly. The amount of nitrous oxide
taken up by blood depends on the amount of blood available: perfusion limited. The time course of O 2
transfer is in between. It does bind to Hb but nothing like the avidity of CO. In normal conditions
capillary PO2 reaches that of alveolar gas when the red cell is about 1/3 of the way along the capillary.

22
Thus, in normal, physiological condition oxygen transfer is perfusion limited. In pathological
conditions, e.g. thickening of alveolar wall, there would be some diffusion limitations as well. PO 2 of
the venous blood and the alveolar air is 40 and 100 mm Hg, respectively. At the end of the capillary
blood PO2 reaches the same value with the alveolar air PO2. During exercise the pulmonary blood flow
is increased and the average travel time of a red blood cell in the capillary is shortened. However, in
normal subjects still there would be no difference between the PO 2 of alveolar air and the blood at the
end of the capillary. On the other hand if there is thickening of alveolar wall oxygen transport would
be impaired and measurable difference between alveolar gas and end-capillary blood PO 2 occurs.
The laws of diffusion state that
VGas = A. D. (P1 – P2)
A: Area
D: Diffusion Constant
Because it is not possible to measure the area and the thickness in living subjects one can introduce D L,
diffusing capacity of the lung.
VGas = DL . (P1 – P2)
DL = VGas / (P1 – P2)
Thus, in normal, physiological condition oxygen transfer is perfusion limited. In pathological
conditions, e.g. thickening of alveolar wall, there would be some diffusion limitations as well. PO 2 of
the venous blood and the alveolar air is 40 and 100 mm Hg, respectively. At the end of the capillary
blood PO2 reaches the same value with the alveolar air PO2. During exercise the pulmonary blood flow
is increased and the average travel time of a red blood cell in the capillary is shortened. However, in
normal subjects still there would be no difference between the PO 2 of alveolar air and the blood at the
end of the capillary. On the other hand if there is thickening of alveolar wall oxygen transport would
be impaired and measurable difference between alveolar gas and end-capillary blood PO 2 occurs.
The laws of diffusion state that
VGas = A. D. (P1 – P2)
A: Area
D: Diffusion Constant
Because it is not possible to measure the area and the thickness in living subjects one can introduce D L,
diffusing capacity of the lung.
VGas = DL . (P1 – P2)
DL = VGas / (P1 – P2)
Secure Best Marks with AI Grader
Need help grading? Try our AI Grader for instant feedback on your assignments.

23
Because transfer of CO is entirely diffusion limited it is an ideal gas to use for diffusion capacity
measurements.
DL = VCO / (P1 – P2)
Since CO in the capillary blood is negligible
DL = VCO / (PACO)
The diffusing capacity of the lung for CO is the volume of CO transferred in mm per mm Hg of
alveolar partial pressure. Single breath method: A single inspiration of a dilute mixture of CO is made
and the rate of disappearance of CO from the alveolar gas during a 10 sec breath hold is calculated.
Steady state method: A subject breathes low concentration of CO until steady state is reached. Then
rate of disappearance of CO from alveolar gas is measured for a short period. The normal value is 25
ml/min/mm Hg. With exercise this value increases 2-3 times. Solubility of CO2 is higher and it
diffuses through tissue 20 times faster than oxygen. Therefore, carbon dioxide transfer is mainly
perfusion limited.
5. PERFUSION
The main function of the pulmonary circulation is to bring systemic venous blood into contact with
alveoli for gas exchange. It begins at the main pulmonary artery, which receives the mixed venous
blood pumped by the right ventricle. This artery then branches successively like the system of airways.
Each time the airway branches, the arterial tree branches that the two parallel each other. The
oxygenated blood is collected from the capillary bed by the pulmonary vein, which drains into the left
atrium. In addition, pulmonary vessels protect the body from obstruction of important vessels in other
organs such as renal or cerebral vessels. When air, fat or blood cloths enter the blood stream (e.g.
during surgery or trauma) pulmonary vessels trap this emboli and endothelial cells release fibrinolytic
substances that help dissolve thrombi. The pulmonary circulation serves as a blood reservoir and the
volume in the lung capillaries is approximately equal to the stroke volume of the right heart.
Pulmonary vessels also contribute to the metabolism of vasoactive hormones. For example angiotensin
I is activated and converted to angiotensin II by angiotensin-converting enzyme which is located on
the surface of the endothelial cells of the pulmonary capillaries.
The differences between the pulmonary and the systemic circulation:
1.The pressures in the pulmonary circulation are remarkably low: The pressure in the main pulmonary
artery is 25 mm Hg (systolic) and 8 mm Hg (diastolic), in average 15 mm Hg. This is a very low-
pressure compare to the pressure in aorta, 100 mm Hg.
2. Another striking property of the pulmonary arteries is their exceedingly thin walls. This anatomical
adaptation of the lung is critically important for its function: The lung is required to receive the
Because transfer of CO is entirely diffusion limited it is an ideal gas to use for diffusion capacity
measurements.
DL = VCO / (P1 – P2)
Since CO in the capillary blood is negligible
DL = VCO / (PACO)
The diffusing capacity of the lung for CO is the volume of CO transferred in mm per mm Hg of
alveolar partial pressure. Single breath method: A single inspiration of a dilute mixture of CO is made
and the rate of disappearance of CO from the alveolar gas during a 10 sec breath hold is calculated.
Steady state method: A subject breathes low concentration of CO until steady state is reached. Then
rate of disappearance of CO from alveolar gas is measured for a short period. The normal value is 25
ml/min/mm Hg. With exercise this value increases 2-3 times. Solubility of CO2 is higher and it
diffuses through tissue 20 times faster than oxygen. Therefore, carbon dioxide transfer is mainly
perfusion limited.
5. PERFUSION
The main function of the pulmonary circulation is to bring systemic venous blood into contact with
alveoli for gas exchange. It begins at the main pulmonary artery, which receives the mixed venous
blood pumped by the right ventricle. This artery then branches successively like the system of airways.
Each time the airway branches, the arterial tree branches that the two parallel each other. The
oxygenated blood is collected from the capillary bed by the pulmonary vein, which drains into the left
atrium. In addition, pulmonary vessels protect the body from obstruction of important vessels in other
organs such as renal or cerebral vessels. When air, fat or blood cloths enter the blood stream (e.g.
during surgery or trauma) pulmonary vessels trap this emboli and endothelial cells release fibrinolytic
substances that help dissolve thrombi. The pulmonary circulation serves as a blood reservoir and the
volume in the lung capillaries is approximately equal to the stroke volume of the right heart.
Pulmonary vessels also contribute to the metabolism of vasoactive hormones. For example angiotensin
I is activated and converted to angiotensin II by angiotensin-converting enzyme which is located on
the surface of the endothelial cells of the pulmonary capillaries.
The differences between the pulmonary and the systemic circulation:
1.The pressures in the pulmonary circulation are remarkably low: The pressure in the main pulmonary
artery is 25 mm Hg (systolic) and 8 mm Hg (diastolic), in average 15 mm Hg. This is a very low-
pressure compare to the pressure in aorta, 100 mm Hg.
2. Another striking property of the pulmonary arteries is their exceedingly thin walls. This anatomical
adaptation of the lung is critically important for its function: The lung is required to receive the

24
whole of the cardiac output at all times. Keeping the pulmonary pressure as low as possible allows
the right heart answer this demand with a minimum work.
3. Unlike the systemic capillaries, which are organised as tubular network with some
interconnections, the pulmonary capillaries mesh together in the alveolar wall so the blood flows as
a thin sheet (capillary bed).
4. Another unique property of the pulmonary circulation is its ability to decrease resistance as cardiac
output increases. Two mechanisms are responsible for this function.
1. Capillary recruitment: opening of initially closed capillaries when cardiac output increases.
2. Capillary distension: The decrease in pulmonary pressure with increased cardiac output has
several beneficial effects: It (1) minimise the load on the right heart, (2) prevents pulmonary
oedema, (3) maintains the adequate flow rate of the blood in the capillary and (4) increases the
capillary surface area.
7. GAS TRANSPORT TO THE PERIPHERY (How do gases move to the
peripheral tissues?)
OXYGEN:
Oxygen is carried in the blood in two forms, dissolved and combined with haemoglobin (Hb).
Dissolved Oxygen: The amount of oxygen dissolved in the blood is proportional to its partial pressure
(Henry’s Law). 100 ml of arterial blood with normal oxygen partial pressure (100 mm Hg) contains
0.3 ml oxygen. By this way amount of oxygen delivered to the tissues is only about 90 ml/min. Taking
whole of the cardiac output at all times. Keeping the pulmonary pressure as low as possible allows
the right heart answer this demand with a minimum work.
3. Unlike the systemic capillaries, which are organised as tubular network with some
interconnections, the pulmonary capillaries mesh together in the alveolar wall so the blood flows as
a thin sheet (capillary bed).
4. Another unique property of the pulmonary circulation is its ability to decrease resistance as cardiac
output increases. Two mechanisms are responsible for this function.
1. Capillary recruitment: opening of initially closed capillaries when cardiac output increases.
2. Capillary distension: The decrease in pulmonary pressure with increased cardiac output has
several beneficial effects: It (1) minimise the load on the right heart, (2) prevents pulmonary
oedema, (3) maintains the adequate flow rate of the blood in the capillary and (4) increases the
capillary surface area.
7. GAS TRANSPORT TO THE PERIPHERY (How do gases move to the
peripheral tissues?)
OXYGEN:
Oxygen is carried in the blood in two forms, dissolved and combined with haemoglobin (Hb).
Dissolved Oxygen: The amount of oxygen dissolved in the blood is proportional to its partial pressure
(Henry’s Law). 100 ml of arterial blood with normal oxygen partial pressure (100 mm Hg) contains
0.3 ml oxygen. By this way amount of oxygen delivered to the tissues is only about 90 ml/min. Taking

25
in to account that the tissue requirements are about 3000 ml Oxygen/min, it is obvious that this way of
transporting oxygen is not adequate for human.
HAEMOGLOBIN:
Haemoglobin (Hb) = Heme (iron-porphyrin) + globin (protein)
Globin has 4 protein polypeptide chains: 2 alpha (each has 141 aa) and 2 beta (each has 146 aa).
Differences in the amino acid sequence of these chains give rise to various types of Hb.
Hb-A: Normal adult Hb
Hb-F: Foetal Hb, which makes part of the total Hb at birth and is gradually, replaced by Hb-A.
Hb-S: S stands for sickle. This Hb has valine in the beta chain instead of glutamic acid. Deoxygenated
form of this Hb is poorly soluble and crystallises in the erythrocytes which results in changes in red
cell shape (crescent or sickle shaped red cells are seen in the blood stream). The fragility of the red
cells is increased and there is a tendency to thrombus formation. Each polypeptide chain is combined
with one heme group. In the centre of each heme group there is one atom of iron, which can combine
with one oxygen molecule. Thus one Hb molecule can bind 4 oxygen molecules. Heme contains iron
in the reduced form (Fe++, ferrous iron). In this form the iron can share electrons and bond with
oxygen. Oxygen forms a reversible combination with Hb.
O2 + Hb ↔ HbO2 (oxyhemoglobin)
in to account that the tissue requirements are about 3000 ml Oxygen/min, it is obvious that this way of
transporting oxygen is not adequate for human.
HAEMOGLOBIN:
Haemoglobin (Hb) = Heme (iron-porphyrin) + globin (protein)
Globin has 4 protein polypeptide chains: 2 alpha (each has 141 aa) and 2 beta (each has 146 aa).
Differences in the amino acid sequence of these chains give rise to various types of Hb.
Hb-A: Normal adult Hb
Hb-F: Foetal Hb, which makes part of the total Hb at birth and is gradually, replaced by Hb-A.
Hb-S: S stands for sickle. This Hb has valine in the beta chain instead of glutamic acid. Deoxygenated
form of this Hb is poorly soluble and crystallises in the erythrocytes which results in changes in red
cell shape (crescent or sickle shaped red cells are seen in the blood stream). The fragility of the red
cells is increased and there is a tendency to thrombus formation. Each polypeptide chain is combined
with one heme group. In the centre of each heme group there is one atom of iron, which can combine
with one oxygen molecule. Thus one Hb molecule can bind 4 oxygen molecules. Heme contains iron
in the reduced form (Fe++, ferrous iron). In this form the iron can share electrons and bond with
oxygen. Oxygen forms a reversible combination with Hb.
O2 + Hb ↔ HbO2 (oxyhemoglobin)
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

26
When oxyhemoglobin dissociates to release oxygen to the tissues (the heme iron is still in ferrous
form) and the Hb is called deoxyhemoglobin (reduced Hb). Oxyhemoglobin is not same with oxidised
Hb (or methemoglobin) in which iron is in the oxidised (Fe +++, ferric) form. Because methemoglobin
lacks the electron necessary to bind oxygen, it does not participate in oxygen transport.
The oxygen carrying capacity of the blood is determined by the Hb concentration. If it is below
normal, anaemia, the oxygen concentration of the blood is reduced. When the Hb concentration is
high, polycythemia, the oxygen carrying capacity of the blood is increased.
The Hb and red blood cell production in the body is under control of erythropoietin, which is produced
by the kidneys. Its production is stimulated when the amount of oxygen delivered to the kidneys is
lower than normal. Normally Hb concentration in men is higher then women, because the red cell
production is also stimulated by androgen.
The oxygen saturation of Hb
O2 combined with Hb / O2 capacity
One gram of Hb can combine with 1.39 ml oxygen and because normal blood has 15 mg of Hb/100 ml
and the oxygen capacity of the 100 ml blood is 20.8 ml.
Oxygen saturation of the arterial blood (PO 2=100 mm Hg) is 97.5 % while oxygen saturation of the
venous blood (PO2= 40 mm Hg) is 75 %.
When oxyhemoglobin dissociates to release oxygen to the tissues (the heme iron is still in ferrous
form) and the Hb is called deoxyhemoglobin (reduced Hb). Oxyhemoglobin is not same with oxidised
Hb (or methemoglobin) in which iron is in the oxidised (Fe +++, ferric) form. Because methemoglobin
lacks the electron necessary to bind oxygen, it does not participate in oxygen transport.
The oxygen carrying capacity of the blood is determined by the Hb concentration. If it is below
normal, anaemia, the oxygen concentration of the blood is reduced. When the Hb concentration is
high, polycythemia, the oxygen carrying capacity of the blood is increased.
The Hb and red blood cell production in the body is under control of erythropoietin, which is produced
by the kidneys. Its production is stimulated when the amount of oxygen delivered to the kidneys is
lower than normal. Normally Hb concentration in men is higher then women, because the red cell
production is also stimulated by androgen.
The oxygen saturation of Hb
O2 combined with Hb / O2 capacity
One gram of Hb can combine with 1.39 ml oxygen and because normal blood has 15 mg of Hb/100 ml
and the oxygen capacity of the 100 ml blood is 20.8 ml.
Oxygen saturation of the arterial blood (PO 2=100 mm Hg) is 97.5 % while oxygen saturation of the
venous blood (PO2= 40 mm Hg) is 75 %.
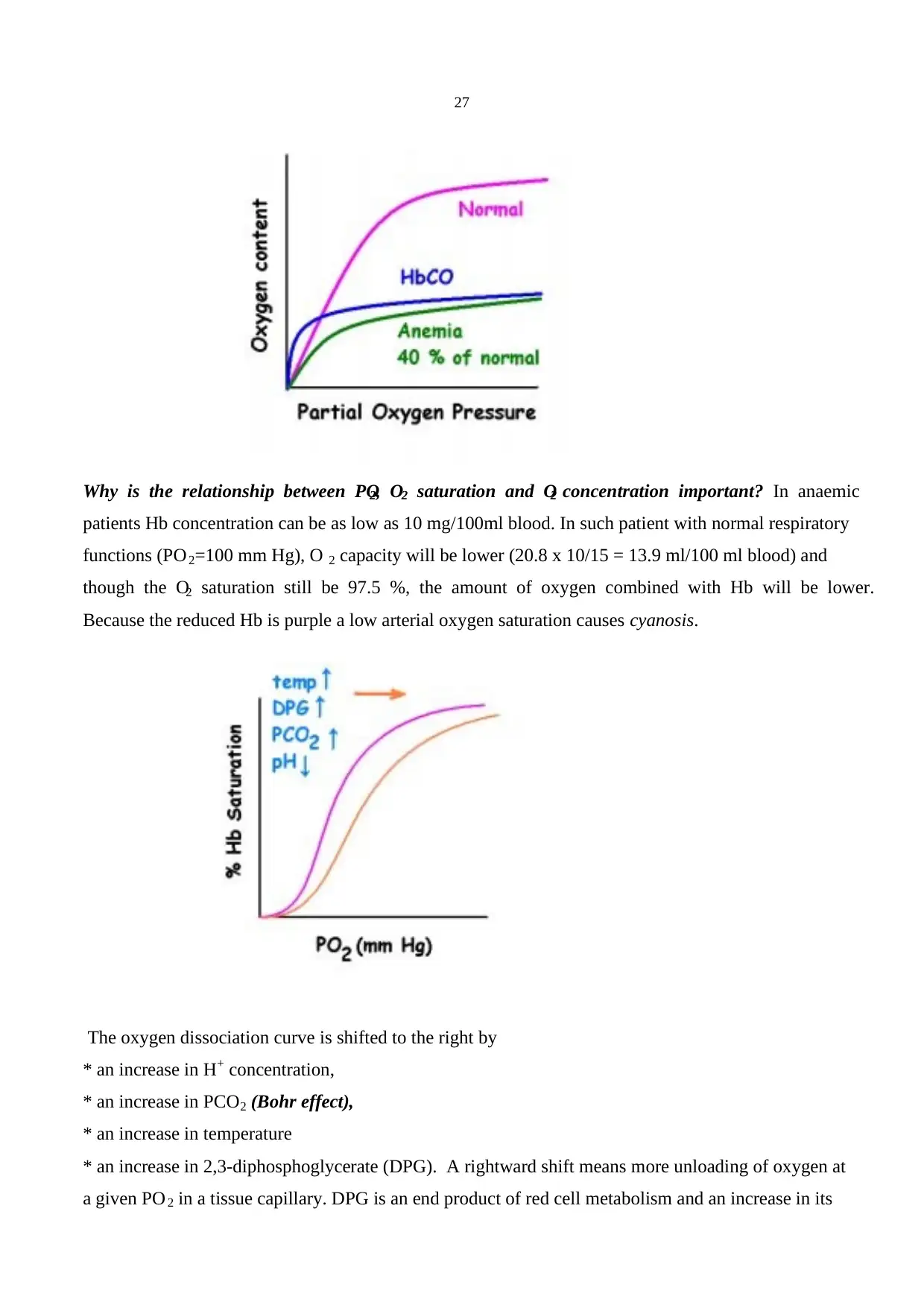
27
Why is the relationship between PO2, O2 saturation and O2 concentration important? In anaemic
patients Hb concentration can be as low as 10 mg/100ml blood. In such patient with normal respiratory
functions (PO2=100 mm Hg), O 2 capacity will be lower (20.8 x 10/15 = 13.9 ml/100 ml blood) and
though the O2 saturation still be 97.5 %, the amount of oxygen combined with Hb will be lower.
Because the reduced Hb is purple a low arterial oxygen saturation causes cyanosis.
The oxygen dissociation curve is shifted to the right by
* an increase in H+ concentration,
* an increase in PCO2 (Bohr effect),
* an increase in temperature
* an increase in 2,3-diphosphoglycerate (DPG). A rightward shift means more unloading of oxygen at
a given PO 2 in a tissue capillary. DPG is an end product of red cell metabolism and an increase in its
Why is the relationship between PO2, O2 saturation and O2 concentration important? In anaemic
patients Hb concentration can be as low as 10 mg/100ml blood. In such patient with normal respiratory
functions (PO2=100 mm Hg), O 2 capacity will be lower (20.8 x 10/15 = 13.9 ml/100 ml blood) and
though the O2 saturation still be 97.5 %, the amount of oxygen combined with Hb will be lower.
Because the reduced Hb is purple a low arterial oxygen saturation causes cyanosis.
The oxygen dissociation curve is shifted to the right by
* an increase in H+ concentration,
* an increase in PCO2 (Bohr effect),
* an increase in temperature
* an increase in 2,3-diphosphoglycerate (DPG). A rightward shift means more unloading of oxygen at
a given PO 2 in a tissue capillary. DPG is an end product of red cell metabolism and an increase in its
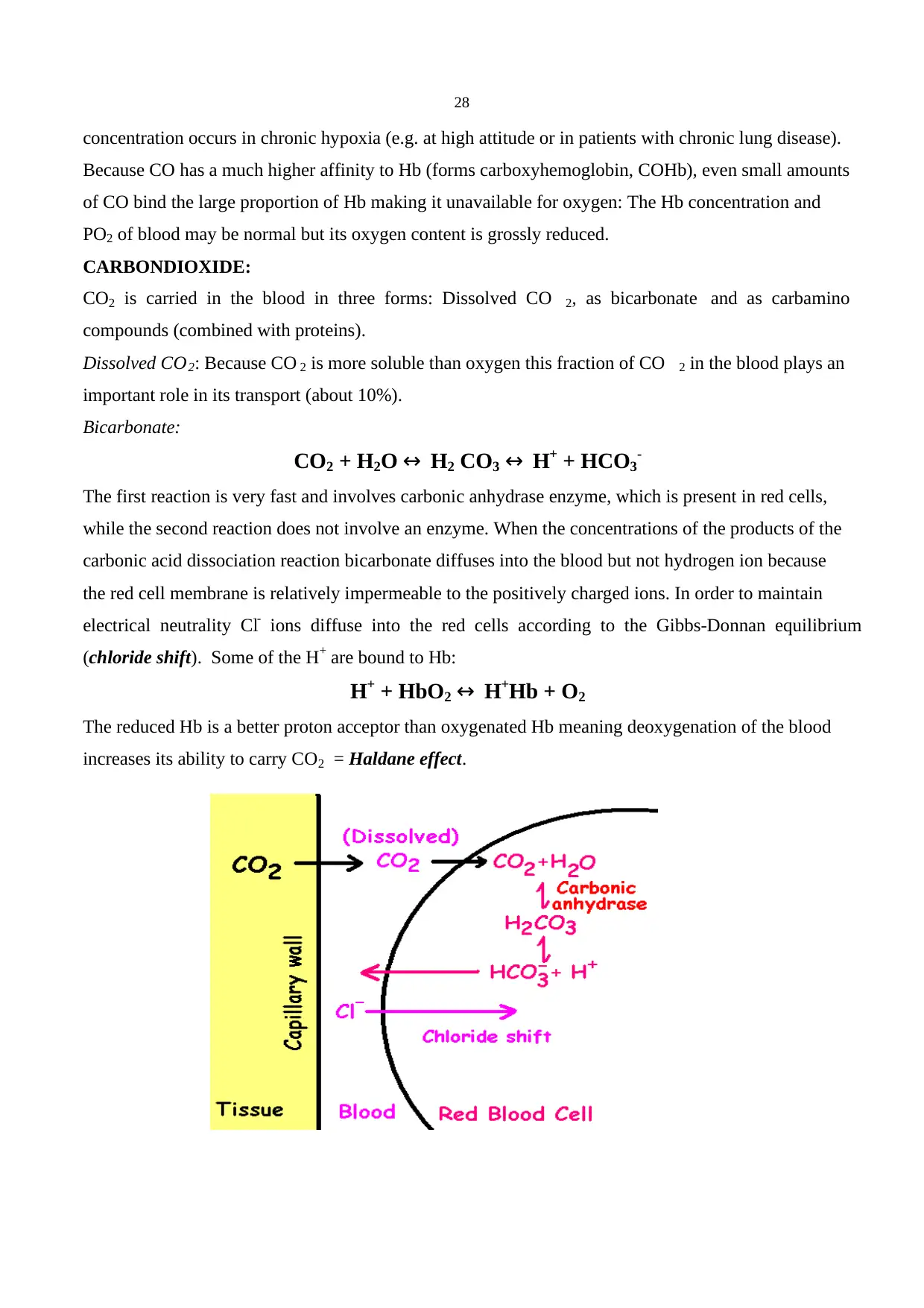
28
concentration occurs in chronic hypoxia (e.g. at high attitude or in patients with chronic lung disease).
Because CO has a much higher affinity to Hb (forms carboxyhemoglobin, COHb), even small amounts
of CO bind the large proportion of Hb making it unavailable for oxygen: The Hb concentration and
PO2 of blood may be normal but its oxygen content is grossly reduced.
CARBONDIOXIDE:
CO2 is carried in the blood in three forms: Dissolved CO 2, as bicarbonate and as carbamino
compounds (combined with proteins).
Dissolved CO2: Because CO 2 is more soluble than oxygen this fraction of CO 2 in the blood plays an
important role in its transport (about 10%).
Bicarbonate:
CO2 + H2O ↔ H2 CO3 ↔ H+ + HCO3-
The first reaction is very fast and involves carbonic anhydrase enzyme, which is present in red cells,
while the second reaction does not involve an enzyme. When the concentrations of the products of the
carbonic acid dissociation reaction bicarbonate diffuses into the blood but not hydrogen ion because
the red cell membrane is relatively impermeable to the positively charged ions. In order to maintain
electrical neutrality Cl- ions diffuse into the red cells according to the Gibbs-Donnan equilibrium
(chloride shift). Some of the H+ are bound to Hb:
H+ + HbO2 ↔ H+Hb + O2
The reduced Hb is a better proton acceptor than oxygenated Hb meaning deoxygenation of the blood
increases its ability to carry CO2 = Haldane effect.
concentration occurs in chronic hypoxia (e.g. at high attitude or in patients with chronic lung disease).
Because CO has a much higher affinity to Hb (forms carboxyhemoglobin, COHb), even small amounts
of CO bind the large proportion of Hb making it unavailable for oxygen: The Hb concentration and
PO2 of blood may be normal but its oxygen content is grossly reduced.
CARBONDIOXIDE:
CO2 is carried in the blood in three forms: Dissolved CO 2, as bicarbonate and as carbamino
compounds (combined with proteins).
Dissolved CO2: Because CO 2 is more soluble than oxygen this fraction of CO 2 in the blood plays an
important role in its transport (about 10%).
Bicarbonate:
CO2 + H2O ↔ H2 CO3 ↔ H+ + HCO3-
The first reaction is very fast and involves carbonic anhydrase enzyme, which is present in red cells,
while the second reaction does not involve an enzyme. When the concentrations of the products of the
carbonic acid dissociation reaction bicarbonate diffuses into the blood but not hydrogen ion because
the red cell membrane is relatively impermeable to the positively charged ions. In order to maintain
electrical neutrality Cl- ions diffuse into the red cells according to the Gibbs-Donnan equilibrium
(chloride shift). Some of the H+ are bound to Hb:
H+ + HbO2 ↔ H+Hb + O2
The reduced Hb is a better proton acceptor than oxygenated Hb meaning deoxygenation of the blood
increases its ability to carry CO2 = Haldane effect.
Secure Best Marks with AI Grader
Need help grading? Try our AI Grader for instant feedback on your assignments.

29
Carbamino compounds: CO2 is also bond by terminal amine groups of several blood proteins. Such as
(the most important one) globin of Hb (carbamino-haemoglobin).
In arterial blood CO 2 is carried 90 % as bicarbonate, 5 % combined with carbamino proteins and 5 %
as dissolved CO2. In venous blood these values are 60, 30 and 10, respectively.
8. ACID-BASE REGULATION
By altering the CO2 elimination the lungs can control the acid-base balance of the body.
H2CO3 ↔ H+ + HCO3-
KA (H2CO3) (H+ + HCO3-)/ H2CO3
KA = (H+ + HCO3-)/ CO2
log KA log H+ + log (HCO3-/ CO2)
-log H+ = -log KA + log (HCO3-/ CO2)
pH = pKA + log (HCO3-/ CO2)
pH = pKA + log (HCO3-/ 0.03 PCO2)
pH = 6.1 + log 24/0.03x40
pH = 6.1 + log 20
pH = 6.1 + 1.3 = 7.4
In the human body bicarbonate concentration is regulated by mainly the kidneys and the PCO 2 by the
lungs.
RESPIRATORY ACIDOSIS: It is due to an increase in PCO2 (e.g. hypoventilation, ventilation-
perfusion inequality). Whenever PCO 2 is increased the bicarbonate concentration is also rises due to
0
20
40
60
80
100
Arterial Venous
Bicarbonate
Carbamino
Dissolved
Carbamino compounds: CO2 is also bond by terminal amine groups of several blood proteins. Such as
(the most important one) globin of Hb (carbamino-haemoglobin).
In arterial blood CO 2 is carried 90 % as bicarbonate, 5 % combined with carbamino proteins and 5 %
as dissolved CO2. In venous blood these values are 60, 30 and 10, respectively.
8. ACID-BASE REGULATION
By altering the CO2 elimination the lungs can control the acid-base balance of the body.
H2CO3 ↔ H+ + HCO3-
KA (H2CO3) (H+ + HCO3-)/ H2CO3
KA = (H+ + HCO3-)/ CO2
log KA log H+ + log (HCO3-/ CO2)
-log H+ = -log KA + log (HCO3-/ CO2)
pH = pKA + log (HCO3-/ CO2)
pH = pKA + log (HCO3-/ 0.03 PCO2)
pH = 6.1 + log 24/0.03x40
pH = 6.1 + log 20
pH = 6.1 + 1.3 = 7.4
In the human body bicarbonate concentration is regulated by mainly the kidneys and the PCO 2 by the
lungs.
RESPIRATORY ACIDOSIS: It is due to an increase in PCO2 (e.g. hypoventilation, ventilation-
perfusion inequality). Whenever PCO 2 is increased the bicarbonate concentration is also rises due to
0
20
40
60
80
100
Arterial Venous
Bicarbonate
Carbamino
Dissolved

30
dissociation of carbonic acid but nevertheless HCO3-/PCO2 falls. In response to these changes the
kidneys start conserving HCO3-.
pH = 6.1 + log 24 / 0.03 x 40 = 6.1 + log20 = 7.4 (Normal)
pH = 6.1 + log 28 / 0.03 x 60 = 6.1 + log15.6 = 7.29 (respiratory acidosis)
pH = 6.1 + log 33 / 0.03 x 60 = 6.1 + log18.3 = 7.36 (compensated res. acidosis)
RESPIRATORY ALKALOSIS: It is caused by a decrease in PCO2 (which in turn increases the
HCO3-/PCO2) due to hyperventilation (e.g. at high attitude, voluntary). The kidneys compensate it by
increasing the HCO3- excretion.
METABOLIC ACIDOSIS: Metabolic refers changes in HCO 3-. In this case HCO 3-/PCO2 decreases
by lowering HCO 3- in the blood. It occurs in conditions accompanied by an accumulation of acids in
the blood (e.g. diabetes mellitus, lactic acid accumulation after tissue hypoxia)
Respiratory compensation occurs by an increase in ventilation that lowers PCO2.
This stimulation is mainly due to stimulation of peripheral chemoreceptors by H+.
METABOLIC ALKALOSIS: An in crease in HCO 3- raises the HCO 3-/PCO2. Common reasons are
excessive ingestion of alkalis and loss of gastric acid due to vomiting. In response the metabolic
acidosis a reduction in alveolar ventilation occurs and PCO2 is increased.
9. RESPIRATORY SYSTEM UNDER STRESS
EXERCISE: During exercise the rate and the depth of breathing are increased. This increase in
ventilation (hyperpnea) matches the simultaneous increase in oxygen consumption and carbon
dioxide production that the arterial blood carbon dioxide and oxygen partial pressures and pH do not
change dramatically (Please note that hyperpnea is different from hyperventilation. In hyperventilation
PCO2 is decreased). The mechanism underlying the exercise-induced changes in ventilation is not
clear.
1. Neurogenic mechanisms: (1) stimulation of respiratory system muscles by sensory nerve activity
from exercising limbs, probably via activating brain stem respiratory centres and/or via spinal reflexes.
(2) stimulatory inputs from cerebral cortex.
2. Chemical mechanisms: Because partial pressures of carbon dioxide and oxygen do not change
during exercise it is difficult to explain possible chemical factors. However, the focal changes in these
parameters near chemoreceptor area may contribute to the exercise-induced changes in ventilation.
Anaerobic threshold: The maximum rate of oxygen consumption than can be attained before blood
lactic acid levels rise as a result of anaerobic respiration. Lactic acid concentration is increased due to
anaerobic limitations in the muscle cells during heavy exercise.
dissociation of carbonic acid but nevertheless HCO3-/PCO2 falls. In response to these changes the
kidneys start conserving HCO3-.
pH = 6.1 + log 24 / 0.03 x 40 = 6.1 + log20 = 7.4 (Normal)
pH = 6.1 + log 28 / 0.03 x 60 = 6.1 + log15.6 = 7.29 (respiratory acidosis)
pH = 6.1 + log 33 / 0.03 x 60 = 6.1 + log18.3 = 7.36 (compensated res. acidosis)
RESPIRATORY ALKALOSIS: It is caused by a decrease in PCO2 (which in turn increases the
HCO3-/PCO2) due to hyperventilation (e.g. at high attitude, voluntary). The kidneys compensate it by
increasing the HCO3- excretion.
METABOLIC ACIDOSIS: Metabolic refers changes in HCO 3-. In this case HCO 3-/PCO2 decreases
by lowering HCO 3- in the blood. It occurs in conditions accompanied by an accumulation of acids in
the blood (e.g. diabetes mellitus, lactic acid accumulation after tissue hypoxia)
Respiratory compensation occurs by an increase in ventilation that lowers PCO2.
This stimulation is mainly due to stimulation of peripheral chemoreceptors by H+.
METABOLIC ALKALOSIS: An in crease in HCO 3- raises the HCO 3-/PCO2. Common reasons are
excessive ingestion of alkalis and loss of gastric acid due to vomiting. In response the metabolic
acidosis a reduction in alveolar ventilation occurs and PCO2 is increased.
9. RESPIRATORY SYSTEM UNDER STRESS
EXERCISE: During exercise the rate and the depth of breathing are increased. This increase in
ventilation (hyperpnea) matches the simultaneous increase in oxygen consumption and carbon
dioxide production that the arterial blood carbon dioxide and oxygen partial pressures and pH do not
change dramatically (Please note that hyperpnea is different from hyperventilation. In hyperventilation
PCO2 is decreased). The mechanism underlying the exercise-induced changes in ventilation is not
clear.
1. Neurogenic mechanisms: (1) stimulation of respiratory system muscles by sensory nerve activity
from exercising limbs, probably via activating brain stem respiratory centres and/or via spinal reflexes.
(2) stimulatory inputs from cerebral cortex.
2. Chemical mechanisms: Because partial pressures of carbon dioxide and oxygen do not change
during exercise it is difficult to explain possible chemical factors. However, the focal changes in these
parameters near chemoreceptor area may contribute to the exercise-induced changes in ventilation.
Anaerobic threshold: The maximum rate of oxygen consumption than can be attained before blood
lactic acid levels rise as a result of anaerobic respiration. Lactic acid concentration is increased due to
anaerobic limitations in the muscle cells during heavy exercise.

31
[The cardiovascular system maintains an adequate amount of oxygen to the tissue that mitochondria can carry out oxidative
phosphorylation. Oxidative phosphorylation is an oxygen consuming process by which energy derived from substrate
oxidation is stored in ATP as a chemical energy. When the oxygen consumption is increased (e.g. heavy exercise) or at
low PO2 (e.g. cellular hypoxia) anaerobic glycolysis becomes a major mechanism for cellular ATP formation and glucose
is reduced to lactate.]
ACCLIMATIZATION TO HIGH ALTITUDE: In the high altitude the human body compensates
the low partial oxygen pressure by changing ventilation or affinity of Hb to oxygen or total Hb
concentration.
Hypoxic ventilatory response: Hyperventilation induced by the decreased partial oxygen pressure.
This lowers the arterial partial carbon dioxide pressure and causes respiratory alkalosis. The rise in
blood pH in turn set the ventilation to a more stable but still slightly higher levels. Hyperventilation
increases the tidal volume and reduces the proportion of the anatomical death space in the inspired air.
This also improves the oxygenation of the blood. However, in spite of all these adaptation
mechanisms, the partial oxygen pressure in the arterial blood can not be increased more than the partial
oxygen pressure in the inspired air. As a result partial pressure of oxygen in the arterial blood
decreases with increasing altitude. At sea levels arterial blood loses 22% of its oxygen load in tissues:
The oxygen saturation of the arterial and venous blood is 97% and 75%, respectively. At high altitude
the low oxygen content of red blood cells stimulates 2-DPG production and decreases the affinity of
Hb to oxygen, which in turn facilitates the oxygen transport to the tissues. However, at very high
altitudes increase in blood pH causes a shift to the left in the oxygen saturation curve and increases the
affinity of Hb to oxygen. This second step is indeed beneficial at very high altitude by increasing the
oxygenation of the blood in the lungs. Due to low oxygen partial pressure in the arterial blood at high
altitude the tissue hypoxia occurs and in response the kidneys secrete erythropoietin hormone.
Erythropoietin stimulates the production of red blood cells resulting in polycythemia, which can cause
oedema, ventricular hypertrophy and heart failure.
[The cardiovascular system maintains an adequate amount of oxygen to the tissue that mitochondria can carry out oxidative
phosphorylation. Oxidative phosphorylation is an oxygen consuming process by which energy derived from substrate
oxidation is stored in ATP as a chemical energy. When the oxygen consumption is increased (e.g. heavy exercise) or at
low PO2 (e.g. cellular hypoxia) anaerobic glycolysis becomes a major mechanism for cellular ATP formation and glucose
is reduced to lactate.]
ACCLIMATIZATION TO HIGH ALTITUDE: In the high altitude the human body compensates
the low partial oxygen pressure by changing ventilation or affinity of Hb to oxygen or total Hb
concentration.
Hypoxic ventilatory response: Hyperventilation induced by the decreased partial oxygen pressure.
This lowers the arterial partial carbon dioxide pressure and causes respiratory alkalosis. The rise in
blood pH in turn set the ventilation to a more stable but still slightly higher levels. Hyperventilation
increases the tidal volume and reduces the proportion of the anatomical death space in the inspired air.
This also improves the oxygenation of the blood. However, in spite of all these adaptation
mechanisms, the partial oxygen pressure in the arterial blood can not be increased more than the partial
oxygen pressure in the inspired air. As a result partial pressure of oxygen in the arterial blood
decreases with increasing altitude. At sea levels arterial blood loses 22% of its oxygen load in tissues:
The oxygen saturation of the arterial and venous blood is 97% and 75%, respectively. At high altitude
the low oxygen content of red blood cells stimulates 2-DPG production and decreases the affinity of
Hb to oxygen, which in turn facilitates the oxygen transport to the tissues. However, at very high
altitudes increase in blood pH causes a shift to the left in the oxygen saturation curve and increases the
affinity of Hb to oxygen. This second step is indeed beneficial at very high altitude by increasing the
oxygenation of the blood in the lungs. Due to low oxygen partial pressure in the arterial blood at high
altitude the tissue hypoxia occurs and in response the kidneys secrete erythropoietin hormone.
Erythropoietin stimulates the production of red blood cells resulting in polycythemia, which can cause
oedema, ventricular hypertrophy and heart failure.
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

32
10. RECOMMENDED FURTHER READING:
1.Anatomy & Physiology by Rod R. Seeley, Trent D. Stephens and Philip Tate McGraw Hill,
International edition.
2. Respiratory Physiology, the essentials by John B West, Williams and Wilkins, (USA).
3. Pulmonary Pathophysiology, the essentials by John B West, Williams and Wilkins, (USA).
4. Principles of Anatomy and Physiology by G.J. Tortora and S.R. Grabowski, Harper Collins College
Publishers (USA).
5. Principles of Physiology by R.M. Berne and M.N. Levy, Mosby (USA).
6. Medical Physiology by R. A. Rhoades & G.A Tanner, Little Brown and Company (USA).
7. Human Anatomy and Physiology by K.M. Van de Graaff and S.R. Fox, Wm. C. Brown Publishers.
10. RECOMMENDED FURTHER READING:
1.Anatomy & Physiology by Rod R. Seeley, Trent D. Stephens and Philip Tate McGraw Hill,
International edition.
2. Respiratory Physiology, the essentials by John B West, Williams and Wilkins, (USA).
3. Pulmonary Pathophysiology, the essentials by John B West, Williams and Wilkins, (USA).
4. Principles of Anatomy and Physiology by G.J. Tortora and S.R. Grabowski, Harper Collins College
Publishers (USA).
5. Principles of Physiology by R.M. Berne and M.N. Levy, Mosby (USA).
6. Medical Physiology by R. A. Rhoades & G.A Tanner, Little Brown and Company (USA).
7. Human Anatomy and Physiology by K.M. Van de Graaff and S.R. Fox, Wm. C. Brown Publishers.

33
11. SELF ASSESSMENT
1. How is a respiratory bronchiole distinguished from a terminal bronchiole?
2. What is the main mechanism of gas movement in the respiratory zone?
3. Which of the following lung volumes cannot be measured with a simple spirometer?
(1) vital capacity,
(2) functional residual capacity,
(3) tidal volume,
(4) residual volume
4. What is pulmonary compliance and which factors may cause changes in it?
5. What is pulmonary sulfactant and what happens in case of its insufficiency?
6. Which factor (PO2, PCO2 or pH) does exert the most important control on ventilation under
physiological conditions and why?
7. What is the Hering-Breuer reflex and its role in normal breathing in adults?
8. Are the neurones in the ventral medullar respiratory centre activated during normal tidal breathing?
9. What is the role of respiratory centres located in the pons in the control of breathing?
10. How does blood pH affects the breathing?
11. What are the main changes occur at high altitudes?
12. How is CO2 is carried in the arterial and venous blood?
13. Patients with ventilation-perfusion inequality have hypoxemia but no CO2 retention. Why?
14. What is chloride shift?
15. What is Haldane effect?
16. What is the difference between that hyperpnea and hyperventilation?
17. What is anaerobic threshold?
11. SELF ASSESSMENT
1. How is a respiratory bronchiole distinguished from a terminal bronchiole?
2. What is the main mechanism of gas movement in the respiratory zone?
3. Which of the following lung volumes cannot be measured with a simple spirometer?
(1) vital capacity,
(2) functional residual capacity,
(3) tidal volume,
(4) residual volume
4. What is pulmonary compliance and which factors may cause changes in it?
5. What is pulmonary sulfactant and what happens in case of its insufficiency?
6. Which factor (PO2, PCO2 or pH) does exert the most important control on ventilation under
physiological conditions and why?
7. What is the Hering-Breuer reflex and its role in normal breathing in adults?
8. Are the neurones in the ventral medullar respiratory centre activated during normal tidal breathing?
9. What is the role of respiratory centres located in the pons in the control of breathing?
10. How does blood pH affects the breathing?
11. What are the main changes occur at high altitudes?
12. How is CO2 is carried in the arterial and venous blood?
13. Patients with ventilation-perfusion inequality have hypoxemia but no CO2 retention. Why?
14. What is chloride shift?
15. What is Haldane effect?
16. What is the difference between that hyperpnea and hyperventilation?
17. What is anaerobic threshold?
1 out of 33
Related Documents






Your All-in-One AI-Powered Toolkit for Academic Success.
+13062052269
info@desklib.com
Available 24*7 on WhatsApp / Email
![[object Object]](/_next/static/media/star-bottom.7253800d.svg)
Unlock your academic potential
© 2024 | Zucol Services PVT LTD | All rights reserved.