BIOL214 Scientific Report: Spectroscopic Analysis of TCA Cycle Enzymes
VerifiedAdded on 2023/06/07
|11
|2156
|435
Report
AI Summary
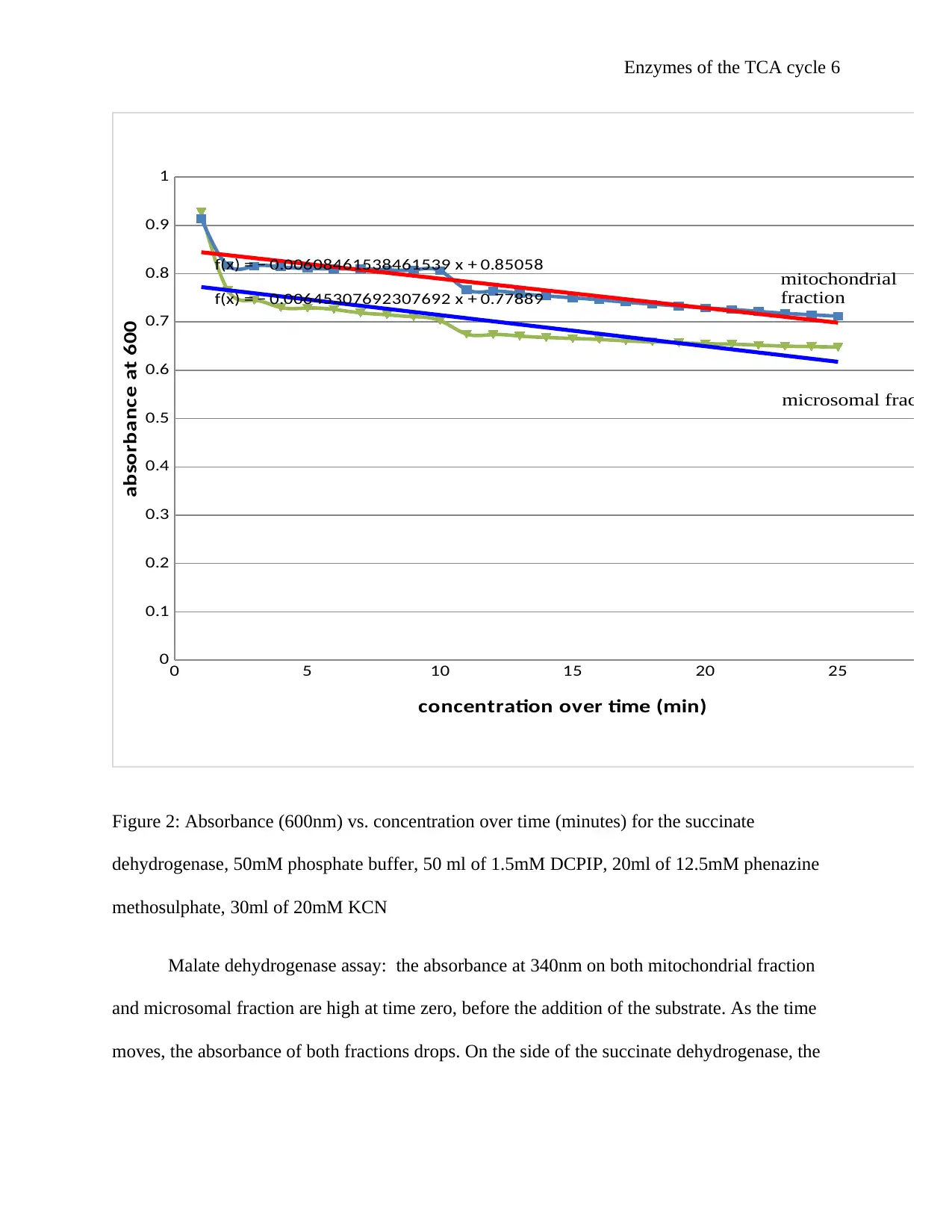
This report details an experiment using spectroscopic methods to analyze the activity of malate dehydrogenase and succinate dehydrogenase, key enzymes in the Tricarboxylic Acid (TCA) cycle. The experiment measured the absorbance of these enzymes in mitochondrial and microsomal fractions of yeast cells, correlating the rate of NADH oxidation with reaction rates. Results, presented as absorbance versus time graphs, indicated enzyme activity in both fractions. The discussion interprets these findings in the context of existing literature, emphasizing the role of DCPIP and PMS in the succinate dehydrogenase assay and explaining the principles behind the spectrophotometric measurements. The report concludes by highlighting the importance of enzyme localization in cellular metabolism and the potential sources of error in the experimental procedure, emphasizing the need for precise dilutions and temperature control. Desklib provides access to similar reports and solved assignments for students.









1 out of 11
Related Documents






Your All-in-One AI-Powered Toolkit for Academic Success.
+13062052269
info@desklib.com
Available 24*7 on WhatsApp / Email
![[object Object]](/_next/static/media/star-bottom.7253800d.svg)
Copyright © 2020–2026 A2Z Services. All Rights Reserved. Developed and managed by ZUCOL.