Review of Recent Advances in Pathogenic E. coli Adherence and Invasion
VerifiedAdded on 2022/08/17
|12
|6941
|14
Report
AI Summary
This report, based on a NIH Public Access Author Manuscript, reviews recent progress in understanding the mechanisms of adherence and invasion used by pathogenic Escherichia coli (E. coli). The review focuses on intestinal and extra-intestinal E. coli strains, detailing how they colonize host cells. It covers enterohemorrhagic E. coli (EHEC), enteropathogenic E. coli (EPEC), enterotoxigenic E. coli (ETEC), and enteroaggregative E. coli (EAEC), highlighting the specific adhesins and invasins each strain utilizes. The report examines the influence of dietary and environmental factors on colonization, including the impact of fiber, diet, and host signaling pathways. It also discusses the role of the type III secretion system (TTSS) and various virulence factors in E. coli pathogenesis. The review emphasizes the need for further research to develop new strategies to combat these infections, focusing on the complex interactions between the bacteria and host epithelial cells. The report is a valuable resource for understanding the complexities of E. coli infections and the ongoing efforts to mitigate their impact.

Recent advances in adherence and invasion of pathogenic
Escherichia coli
Anjana Kalita1,*, Jia Hu1,*, and Alfredo G. Torres1,2,†
1Department of Microbiology and Immunology, University of Texas Medical Branch, Galveston,
Texas, 77555, USA
2Department of Pathology, University of Texas Medical Branch, Galveston, Texas, 77555, USA
Abstract
Purpose of review—Colonization of the host epithelia by pathogenic Escherichia coli is
influenced by the ability of the bacteria to interact with host surfaces. Because the initial step of an
E. coli infection is to adhere, invade, and persist within host cells, some strategies used by
intestinal and extra-intestinal E. coli to infect host cell are presented.
Recent findings—This review highlights recent progress understanding how extra-intestinal
pathogenic E. coli strains express specific adhesins/invasins that allow colonization of the urinary
tract or the meninges, while intestinal E. coli strains are able to colonize different regions of the
intestinal tract using other specialized adhesins/invasins. Finally, evaluation of, different diets and
environmental conditions regulating the colonization of these pathogens is discussed.
Summary—Discovery of new interactions between pathogenic E. coli and the host epithelial
cells unravels the need of more mechanistic studies that can provide new clues in how to combat
these infections.
Keywords
enterohemorrhagic E. coli; enteropathogenic; enterotoxigenic; uropathogenic; enteroaggregative;
adherent invasive E. coli
Introduction
Escherichia coli are commonly found as part of the gut flora, where it is the predominant
aerobic organism, living in symbiosis with its vertebrate host. However, there are several
categories of E. coli strains that have acquired the ability to cause pathogenic processes in
the host (1). These E. coli strains can cause intestinal (enteritis, diarrhea, or dysentery), or.
extra-intestinal diseases (urinary tract infections, sepsis, or meningitis) (2, 3). To cause
infection, pathogenic E. coli interact with the mucosa, by either attaching to the epithelial
†Corresponding author: Alfredo G. Torres, PhD. UTMB, Department of Microbiology and Immunology, Galveston, Texas, 77555
Phone (409) 747 0189; Fax (409) 747 6869; altorres@utmb.edu.
*These authors contributed equally to this work
Conflicts of interest
There are no conflicts of interest.
NIH Public Access
Author Manuscript
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
Published in final edited form as:
Curr Opin Infect Dis. 2014 October ; 27(5): 459–464. doi:10.1097/QCO.0000000000000092.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Escherichia coli
Anjana Kalita1,*, Jia Hu1,*, and Alfredo G. Torres1,2,†
1Department of Microbiology and Immunology, University of Texas Medical Branch, Galveston,
Texas, 77555, USA
2Department of Pathology, University of Texas Medical Branch, Galveston, Texas, 77555, USA
Abstract
Purpose of review—Colonization of the host epithelia by pathogenic Escherichia coli is
influenced by the ability of the bacteria to interact with host surfaces. Because the initial step of an
E. coli infection is to adhere, invade, and persist within host cells, some strategies used by
intestinal and extra-intestinal E. coli to infect host cell are presented.
Recent findings—This review highlights recent progress understanding how extra-intestinal
pathogenic E. coli strains express specific adhesins/invasins that allow colonization of the urinary
tract or the meninges, while intestinal E. coli strains are able to colonize different regions of the
intestinal tract using other specialized adhesins/invasins. Finally, evaluation of, different diets and
environmental conditions regulating the colonization of these pathogens is discussed.
Summary—Discovery of new interactions between pathogenic E. coli and the host epithelial
cells unravels the need of more mechanistic studies that can provide new clues in how to combat
these infections.
Keywords
enterohemorrhagic E. coli; enteropathogenic; enterotoxigenic; uropathogenic; enteroaggregative;
adherent invasive E. coli
Introduction
Escherichia coli are commonly found as part of the gut flora, where it is the predominant
aerobic organism, living in symbiosis with its vertebrate host. However, there are several
categories of E. coli strains that have acquired the ability to cause pathogenic processes in
the host (1). These E. coli strains can cause intestinal (enteritis, diarrhea, or dysentery), or.
extra-intestinal diseases (urinary tract infections, sepsis, or meningitis) (2, 3). To cause
infection, pathogenic E. coli interact with the mucosa, by either attaching to the epithelial
†Corresponding author: Alfredo G. Torres, PhD. UTMB, Department of Microbiology and Immunology, Galveston, Texas, 77555
Phone (409) 747 0189; Fax (409) 747 6869; altorres@utmb.edu.
*These authors contributed equally to this work
Conflicts of interest
There are no conflicts of interest.
NIH Public Access
Author Manuscript
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
Published in final edited form as:
Curr Opin Infect Dis. 2014 October ; 27(5): 459–464. doi:10.1097/QCO.0000000000000092.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

cells and in some instances, invading the target host cells. Because bacterial adhesion and/or
invasion to/into host cells are the first step during infection, it is necessary to understand at a
molecular level the mechanisms mediating these initial interactions. This article focus on
reviewing recent progress on the understanding of the adhesion/invasion mechanisms used
by intestinal and extra-intestinal pathogenic E. coli during colonization of the host cells.
Enterohemorrhagic E. coli (EHEC)
EHEC are a category of pathogenic E. coli that colonize the human large intestine and which
can cause bloody diarrhea, or a systemic process known as hemolytic uremic syndrome (4).
EHEC strains are characterized by the production of Shiga toxin and the formation of
attaching and effacing intestinal lesions (Figure 1). Cattle are a main reservoir for EHEC
strains; however several vegetables and fruits can serve as vehicles for EHEC outbreaks (5).
EHEC colonization is impacted by nutrient availability and dietary choice. Zumbrun et al
found that dietary fiber content affects susceptibility to E. coli O157:H7 infection in mice
(6). They treated BALB/c mice with high fiber diet (10% guar gum) or low fiber diet (2%
guar gum) for two weeks and then mice were challenge with 109 to 1011 cfu of E. coli
O157:H7. The results showed that mice fed with high fiber diet had enhanced levels of
butyrate that temporally increased the expression of the Shiga toxin receptor Gb3.
Therefore, mice exhibited greater E. coli O157:H7 colonization and reduction in resident
Escherichia spp. Sheng et al also showed that cattle fed a hay diet are colonized by EHEC
for a longer period of time than grain fed cattle (7). Different diets regulate the colonization
of E. coli O157:H7 by altering the composition of gastrointestinal tract microbiota and the
study demonstrated that the bacterial SdiA sensor activates genes conferring EHEC acid
resistance, increasing efficient colonization of the cattle mucosa (8).
Modulation of host signals in the intestinal epithelia also affects EHEC colonization.
Intestinal epithelial cells produced SIGIRR, a negative regulator of interleukin (IL)-1 and
TLR signaling, that makes the cells hypo-responsive (9, 10). To address whether hypo-
responsiveness affects enteric host defense, Sham et al challenge Sigirr deficient (−/−) mice
with the murine pathogen, EHEC-related, Citrobacter rodentium and showed that Sigirr−/−
mice are more susceptible to bacterial infection and had a dramatic loss of microbiota (11).
The study showed that this host signaling mechanism promotes commensal dependent
resistance to EHEC colonization.
Type III secretion system (TTSS) is required for EHEC colonization and attaching and
effacing lesion formation. This syringe-like structure used to inject virulence factors into the
host cell is exquisitely regulated. Hansen et al revealed that tyrosine phosphorylation in
EHEC mediates signaling of virulence properties, including the type III secretion system
(12). SspA is a known regulator of the TTSS (13) and a phosphorylated tyrosine residue of
this protein positively affects expression and secretion of type III secretion system proteins.
Branchu et al also found a new regulator of the TTSS (14) known as the NO-sensor
regulator, NsrR. Nitric oxide (NO) reduced EHEC adherence to intestinal epithelial cells, by
causing the detachment of the NsrR activator from the type III secretion system-encoding
operons (LEE1/4/5), limiting colonization.
Kalita et al. Page 2
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
invasion to/into host cells are the first step during infection, it is necessary to understand at a
molecular level the mechanisms mediating these initial interactions. This article focus on
reviewing recent progress on the understanding of the adhesion/invasion mechanisms used
by intestinal and extra-intestinal pathogenic E. coli during colonization of the host cells.
Enterohemorrhagic E. coli (EHEC)
EHEC are a category of pathogenic E. coli that colonize the human large intestine and which
can cause bloody diarrhea, or a systemic process known as hemolytic uremic syndrome (4).
EHEC strains are characterized by the production of Shiga toxin and the formation of
attaching and effacing intestinal lesions (Figure 1). Cattle are a main reservoir for EHEC
strains; however several vegetables and fruits can serve as vehicles for EHEC outbreaks (5).
EHEC colonization is impacted by nutrient availability and dietary choice. Zumbrun et al
found that dietary fiber content affects susceptibility to E. coli O157:H7 infection in mice
(6). They treated BALB/c mice with high fiber diet (10% guar gum) or low fiber diet (2%
guar gum) for two weeks and then mice were challenge with 109 to 1011 cfu of E. coli
O157:H7. The results showed that mice fed with high fiber diet had enhanced levels of
butyrate that temporally increased the expression of the Shiga toxin receptor Gb3.
Therefore, mice exhibited greater E. coli O157:H7 colonization and reduction in resident
Escherichia spp. Sheng et al also showed that cattle fed a hay diet are colonized by EHEC
for a longer period of time than grain fed cattle (7). Different diets regulate the colonization
of E. coli O157:H7 by altering the composition of gastrointestinal tract microbiota and the
study demonstrated that the bacterial SdiA sensor activates genes conferring EHEC acid
resistance, increasing efficient colonization of the cattle mucosa (8).
Modulation of host signals in the intestinal epithelia also affects EHEC colonization.
Intestinal epithelial cells produced SIGIRR, a negative regulator of interleukin (IL)-1 and
TLR signaling, that makes the cells hypo-responsive (9, 10). To address whether hypo-
responsiveness affects enteric host defense, Sham et al challenge Sigirr deficient (−/−) mice
with the murine pathogen, EHEC-related, Citrobacter rodentium and showed that Sigirr−/−
mice are more susceptible to bacterial infection and had a dramatic loss of microbiota (11).
The study showed that this host signaling mechanism promotes commensal dependent
resistance to EHEC colonization.
Type III secretion system (TTSS) is required for EHEC colonization and attaching and
effacing lesion formation. This syringe-like structure used to inject virulence factors into the
host cell is exquisitely regulated. Hansen et al revealed that tyrosine phosphorylation in
EHEC mediates signaling of virulence properties, including the type III secretion system
(12). SspA is a known regulator of the TTSS (13) and a phosphorylated tyrosine residue of
this protein positively affects expression and secretion of type III secretion system proteins.
Branchu et al also found a new regulator of the TTSS (14) known as the NO-sensor
regulator, NsrR. Nitric oxide (NO) reduced EHEC adherence to intestinal epithelial cells, by
causing the detachment of the NsrR activator from the type III secretion system-encoding
operons (LEE1/4/5), limiting colonization.
Kalita et al. Page 2
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Enteropathogenic E. coli (EPEC)
EPEC isolates colonize the small intestine and are one of the leading causes of infantile fatal
diarrhea (15). In industrialized countries, there are sporadic diarrheal cases in daycare
facilities (16, 17). As for EHEC, EPEC uses the type III secretion system to form attaching
and effacing lesions (Figure 1). EPEC strains are subdivided into typical (tEPEC) and
atypical EPEC (aEPEC) based on the presence of EPEC adherence factor plasmid associated
with the tEPEC localized adherence pattern (18). Some aEPEC form diffuse (DA) or
aggregative adherence pattern and Hernandes et al (19) found that the DA pattern is
associated with the TTSS system and its suggested that the traslocon serves as the DA
adhesin.
Regarding the type III secretion system, recent studies have evaluated the role of the effector
protein NleB in pathogenesis. Deletion of nleB1 in C. rodentium caused significant
reduction in murine intestinal colonization (20), and NleB has also been shown to modulate
the host innate immune system by suppressing tumor necrosis factor (TNF)-mediated NF-
κB activation (21). Three studies have investigated the role of NleB1 in EPEC. Gao et al
focused on how NleB interfere with NF-κB (22). A proteomic screen was used to identify
the host GAPDH as NleB-interacting protein, which results in modification of GAPDH and
inhibition of NF-κB-dependent innate immune responses. The other two groups discovered
that NleB blocks host death receptor signaling (23, 24), by interacting with two death
receptor-signaling proteins, TRADD and FADD. NleB is the first known bacterial virulence
factor to target death receptor signaling and it has been suggested that blocking this
signaling mechanism may facilitate EPEC and EHEC colonization.
Other studies evaluated ways to reduce EPEC adhesion to host cells. Pan et al expressed
synthetic tetrameric-branched peptide that enhanced the expression of Mucin 3 (25). They
found that Mucin 3 interacted with EPEC and EHEC and reduced their binding to epithelial
cells. In other study, Salcedo et al showed that the combination of gangliosides and sialic
acid were able to interfere with EPEC and EHEC adhesion to Caco-2 cells (26).
Enterotoxigenic E. coli (ETEC)
ETEC colonizes the human small intestine and is responsible for neonatal diarrhea in
developing countries as well as “travelers’ diarrhea” (27). ETEC adherence to the intestinal
mucosa is mainly mediated by diverse adhesive structures known as colonization factors,
which in combination with the heat-labile (LT) and/or heat-stable (ST) enterotoxins, causes
disruption of fluid homeostasis in the host, resulting in diarrhea (2, 28) (Figure 1).
Guevara et al recently investigated one colonization factors, the CS21 pilus, and its role in
adherence and pathogenesis in vivo (29). They found that ETEC CS21 (Longus) adherence
to primary intestinal cells was inhibited by anti-LngA sera and the purified LngA protein. In
vivo intra-stomach administration of CS21-expressing ETEC strain contributes to 100%
lethality of newborn mice, which was reduced in the lngA mutant. A separate study
investigated the role of the LT toxin in ETEC colonization (30). The authors observed
enhanced adherence to IPEC-J2 cells by various isogenic ETEC constructs carrying different
forms of the LT toxin (K88/LT wild type, attenuated toxin form [K88/LTR192G] and
Kalita et al. Page 3
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
EPEC isolates colonize the small intestine and are one of the leading causes of infantile fatal
diarrhea (15). In industrialized countries, there are sporadic diarrheal cases in daycare
facilities (16, 17). As for EHEC, EPEC uses the type III secretion system to form attaching
and effacing lesions (Figure 1). EPEC strains are subdivided into typical (tEPEC) and
atypical EPEC (aEPEC) based on the presence of EPEC adherence factor plasmid associated
with the tEPEC localized adherence pattern (18). Some aEPEC form diffuse (DA) or
aggregative adherence pattern and Hernandes et al (19) found that the DA pattern is
associated with the TTSS system and its suggested that the traslocon serves as the DA
adhesin.
Regarding the type III secretion system, recent studies have evaluated the role of the effector
protein NleB in pathogenesis. Deletion of nleB1 in C. rodentium caused significant
reduction in murine intestinal colonization (20), and NleB has also been shown to modulate
the host innate immune system by suppressing tumor necrosis factor (TNF)-mediated NF-
κB activation (21). Three studies have investigated the role of NleB1 in EPEC. Gao et al
focused on how NleB interfere with NF-κB (22). A proteomic screen was used to identify
the host GAPDH as NleB-interacting protein, which results in modification of GAPDH and
inhibition of NF-κB-dependent innate immune responses. The other two groups discovered
that NleB blocks host death receptor signaling (23, 24), by interacting with two death
receptor-signaling proteins, TRADD and FADD. NleB is the first known bacterial virulence
factor to target death receptor signaling and it has been suggested that blocking this
signaling mechanism may facilitate EPEC and EHEC colonization.
Other studies evaluated ways to reduce EPEC adhesion to host cells. Pan et al expressed
synthetic tetrameric-branched peptide that enhanced the expression of Mucin 3 (25). They
found that Mucin 3 interacted with EPEC and EHEC and reduced their binding to epithelial
cells. In other study, Salcedo et al showed that the combination of gangliosides and sialic
acid were able to interfere with EPEC and EHEC adhesion to Caco-2 cells (26).
Enterotoxigenic E. coli (ETEC)
ETEC colonizes the human small intestine and is responsible for neonatal diarrhea in
developing countries as well as “travelers’ diarrhea” (27). ETEC adherence to the intestinal
mucosa is mainly mediated by diverse adhesive structures known as colonization factors,
which in combination with the heat-labile (LT) and/or heat-stable (ST) enterotoxins, causes
disruption of fluid homeostasis in the host, resulting in diarrhea (2, 28) (Figure 1).
Guevara et al recently investigated one colonization factors, the CS21 pilus, and its role in
adherence and pathogenesis in vivo (29). They found that ETEC CS21 (Longus) adherence
to primary intestinal cells was inhibited by anti-LngA sera and the purified LngA protein. In
vivo intra-stomach administration of CS21-expressing ETEC strain contributes to 100%
lethality of newborn mice, which was reduced in the lngA mutant. A separate study
investigated the role of the LT toxin in ETEC colonization (30). The authors observed
enhanced adherence to IPEC-J2 cells by various isogenic ETEC constructs carrying different
forms of the LT toxin (K88/LT wild type, attenuated toxin form [K88/LTR192G] and
Kalita et al. Page 3
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
⊘ This is a preview!⊘
Do you want full access?
Subscribe today to unlock all pages.
Trusted by 1+ million students worldwide

expressing just the B subunit [K88/LTB]), in contrast to the attenuated phenotype of the LT-
negative construct. LT+ strains blocking binding of wild type ETEC strain to IPEC-J2 cells
suggested that LT-driven adherence alters net surface charge on epithelial cells. Another
study evaluated the transcriptional pattern of 214 genes at different time points following
interaction of prototype ETEC E24377A with epithelial cells (31). The study found a
prominent alteration of genes associated with motility, adhesion, toxin production, and
global regulatory mechanisms, such as those linked to cAMP receptor protein and c-di-
GMP, upon ETEC-host interaction, which suggested that ETEC coordinated its responses to
the host environment by sequential activation of different virulence factors.
Among those ETEC strains infecting animals, the most common adhesive fimbriae include
K88 or K99 (also called F4 and F5) (28). Recently, Zhou et al have demonstrated that
deletion of fliC (encoding the major flagellin protein) and/or the faeG (encoding the F4
major fimbrial subunit) from ETEC strain C83902 significantly reduced its ability to adhere
to porcine epithelia IPEC-J2 cells, but also impacting biofilm formation and quorum sensing
(32). Interestingly, another study found an alternative way to block the adherence of ETEC
K88 to IPEC-J2 by using ETEC anti-adhesives, including casein glycomacropeptide,
exopolysaccharide, and vegetable extract (locust bean or wheat bran) (33). Finally, studies
with human milk and commercial infant formulas found that the main gangliosides (GM3,
GD3, GM1) and free sialic acid (Neu5Ac) are able to impede the adhesion of several
pathogenic bacteria, including ETEC (26). Other dietary supplements, such as plantain NSP,
also hampered the adherence of ETEC to Caco-2 cells, and has been suggested that blocking
M-cell bacterial translocation can subsequently prevent diarrheal episodes (34).
Enteroaggregative E. coli (EAEC)
EAEC are a major cause of acute and persistent diarrhea in the small intestine of children
and adults worldwide, including industrialized countries (35). EAEC is also responsible for
sporadic cases and several outbreaks (36). At an initial stage of infection, EAEC adhere in a
characteristic “stacked-brick” formation to host intestinal mucosa; forming a thick mucoid
biofilm. The adherence process is mainly mediated by fimbrial structures called aggregative
adherence fimbriae (AAF) (35) (Figure 1). Additionally, several EAEC virulence-related
genes have been described but their role in the clinical outcome of infection is not
completely defined.
A study recently confirmed a high prevalence, endemicity and heterogeneity of EAEC
strains, and found that the plasmid-encoded toxin or AAF/II fimbrial subunit genes were
associated significantly with disease (37). However, this study also demonstrated that the
pathophysiology of EAEC infections involves a complex and dynamic modulation of several
virulence factors. Another study identified an association of the EAEC virulence-encoded
aggR gene (virulence regulator), pCDV432 plasmid, and additional virulence gene products,
including dispersin and the Air adhesin in 90% of the diarrheagenic isolates that
distinguished them from non-diarrheagenic EAEC strains, suggesting heterogeneity among
highly pathogenic EAEC strains (38).
Kalita et al. Page 4
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
negative construct. LT+ strains blocking binding of wild type ETEC strain to IPEC-J2 cells
suggested that LT-driven adherence alters net surface charge on epithelial cells. Another
study evaluated the transcriptional pattern of 214 genes at different time points following
interaction of prototype ETEC E24377A with epithelial cells (31). The study found a
prominent alteration of genes associated with motility, adhesion, toxin production, and
global regulatory mechanisms, such as those linked to cAMP receptor protein and c-di-
GMP, upon ETEC-host interaction, which suggested that ETEC coordinated its responses to
the host environment by sequential activation of different virulence factors.
Among those ETEC strains infecting animals, the most common adhesive fimbriae include
K88 or K99 (also called F4 and F5) (28). Recently, Zhou et al have demonstrated that
deletion of fliC (encoding the major flagellin protein) and/or the faeG (encoding the F4
major fimbrial subunit) from ETEC strain C83902 significantly reduced its ability to adhere
to porcine epithelia IPEC-J2 cells, but also impacting biofilm formation and quorum sensing
(32). Interestingly, another study found an alternative way to block the adherence of ETEC
K88 to IPEC-J2 by using ETEC anti-adhesives, including casein glycomacropeptide,
exopolysaccharide, and vegetable extract (locust bean or wheat bran) (33). Finally, studies
with human milk and commercial infant formulas found that the main gangliosides (GM3,
GD3, GM1) and free sialic acid (Neu5Ac) are able to impede the adhesion of several
pathogenic bacteria, including ETEC (26). Other dietary supplements, such as plantain NSP,
also hampered the adherence of ETEC to Caco-2 cells, and has been suggested that blocking
M-cell bacterial translocation can subsequently prevent diarrheal episodes (34).
Enteroaggregative E. coli (EAEC)
EAEC are a major cause of acute and persistent diarrhea in the small intestine of children
and adults worldwide, including industrialized countries (35). EAEC is also responsible for
sporadic cases and several outbreaks (36). At an initial stage of infection, EAEC adhere in a
characteristic “stacked-brick” formation to host intestinal mucosa; forming a thick mucoid
biofilm. The adherence process is mainly mediated by fimbrial structures called aggregative
adherence fimbriae (AAF) (35) (Figure 1). Additionally, several EAEC virulence-related
genes have been described but their role in the clinical outcome of infection is not
completely defined.
A study recently confirmed a high prevalence, endemicity and heterogeneity of EAEC
strains, and found that the plasmid-encoded toxin or AAF/II fimbrial subunit genes were
associated significantly with disease (37). However, this study also demonstrated that the
pathophysiology of EAEC infections involves a complex and dynamic modulation of several
virulence factors. Another study identified an association of the EAEC virulence-encoded
aggR gene (virulence regulator), pCDV432 plasmid, and additional virulence gene products,
including dispersin and the Air adhesin in 90% of the diarrheagenic isolates that
distinguished them from non-diarrheagenic EAEC strains, suggesting heterogeneity among
highly pathogenic EAEC strains (38).
Kalita et al. Page 4
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

Another area of study in EAEC pathogenesis is the contribution of the autotransporter
proteins. Munera et al evaluated the role of chromosome-encoded autotransporters in
colonization and subsequent induction of diarrheal disease in infant rabbits and found that
Shiga toxin-producing EAEC O104:H4 autotransporters, but not its virulence plasmid, are
critical for robust colonization and disease (39). EAEC has also been associated with urinary
tract infections (40, 41). The autotrasporter Pic has been defined as a gene marker associated
with spreading of infection to the urinary tract (40). Finally, comparison of EAEC isolates
from HIV-positive and non-HIV diarrheal samples showed that HIV-positive isolates are
stronger biofilm producers and more resistant to antibiotics than the non-HIV diarrheal
isolates, which confirmed the heterogeneity of the EAEC isolates (42).
Adherent and Invasive E. coli (AIEC)
Inflammatory bowel disease (IBD), particularly Crohn’s disease and ulcerative colitis, are
the result of alterations in the intestinal microbiota due to a variety of genetic and
environmental factors (43). Interestingly, an increased number of AIEC have been isolated
from IBD patients and more frequently found in ileal-Crohn’s disease patients than in
healthy controls (44). AIEC strains have the ability to adhere and invade intestinal epithelial
cells and survive within macrophages (45). Small et al recently established a chronic
infection murine model using prototypical AIEC isolates (46), and demonstrated that AIEC
infection stimulates chronic inflammation and fibrosis in mice. This study showed for the
first time evidence that an infection with AIEC can cause intestinal symptoms similar to
those observed in Crohn’s disease patients.
AIEC adherence depends on the expression of the type 1 pili, long polar fimbriae and the
presence of the carcinoembryonic antigen (CEACAM6) as a host cell receptor (47, 48)
(Figure 1). Crohn’s disease patients showed abnormal expression of CEACAM6 (48) and as
such, Martinez-Medina et al used a model of transgenic mice expressing CEACAMs to
assess the effects of a high fat/high sugar western diet on gut microbiota composition,
barrier integrity and susceptibility to infection (49). They found that the diet induces
changes in gut microbiota composition, altering host homeostasis and promoting AIEC
murine gut colonization. With respect to the type 1 pili, a study sequenced the fimH gene
(FimH is the adhesin protein located on the tip of the pili) from 45 AIEC strains and 47 non-
AIEC E. coli strains. Phylogenetic analysis found that AIEC strains predominantly express
FimH with amino acid mutations of a recent evolutionary origin as compared to non-AIEC
strains, which represents a feature of pathoadaptive changes in several bacterial pathogens
(50). The accumulation of these mutations confers AIEC the ability to adhere to CEACAM-
expressing intestinal epithelial cells and to participate in the development of chronic
inflammation in a genetically susceptible host. Finally, it is known that the long polar
fimbriae help AIEC to interact with intestinal Peyer’s patches and M cells (51). A recent
study analyzed the effect of gastrointestinal conditions on AIEC long polar fimbriae
expression. The authors found that bile salts strongly enhanced fimbriae expression, causing
a higher level of interaction of AIEC with Peyer’s patches and a higher level of translocation
through M cell monolayers (52).
Kalita et al. Page 5
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
proteins. Munera et al evaluated the role of chromosome-encoded autotransporters in
colonization and subsequent induction of diarrheal disease in infant rabbits and found that
Shiga toxin-producing EAEC O104:H4 autotransporters, but not its virulence plasmid, are
critical for robust colonization and disease (39). EAEC has also been associated with urinary
tract infections (40, 41). The autotrasporter Pic has been defined as a gene marker associated
with spreading of infection to the urinary tract (40). Finally, comparison of EAEC isolates
from HIV-positive and non-HIV diarrheal samples showed that HIV-positive isolates are
stronger biofilm producers and more resistant to antibiotics than the non-HIV diarrheal
isolates, which confirmed the heterogeneity of the EAEC isolates (42).
Adherent and Invasive E. coli (AIEC)
Inflammatory bowel disease (IBD), particularly Crohn’s disease and ulcerative colitis, are
the result of alterations in the intestinal microbiota due to a variety of genetic and
environmental factors (43). Interestingly, an increased number of AIEC have been isolated
from IBD patients and more frequently found in ileal-Crohn’s disease patients than in
healthy controls (44). AIEC strains have the ability to adhere and invade intestinal epithelial
cells and survive within macrophages (45). Small et al recently established a chronic
infection murine model using prototypical AIEC isolates (46), and demonstrated that AIEC
infection stimulates chronic inflammation and fibrosis in mice. This study showed for the
first time evidence that an infection with AIEC can cause intestinal symptoms similar to
those observed in Crohn’s disease patients.
AIEC adherence depends on the expression of the type 1 pili, long polar fimbriae and the
presence of the carcinoembryonic antigen (CEACAM6) as a host cell receptor (47, 48)
(Figure 1). Crohn’s disease patients showed abnormal expression of CEACAM6 (48) and as
such, Martinez-Medina et al used a model of transgenic mice expressing CEACAMs to
assess the effects of a high fat/high sugar western diet on gut microbiota composition,
barrier integrity and susceptibility to infection (49). They found that the diet induces
changes in gut microbiota composition, altering host homeostasis and promoting AIEC
murine gut colonization. With respect to the type 1 pili, a study sequenced the fimH gene
(FimH is the adhesin protein located on the tip of the pili) from 45 AIEC strains and 47 non-
AIEC E. coli strains. Phylogenetic analysis found that AIEC strains predominantly express
FimH with amino acid mutations of a recent evolutionary origin as compared to non-AIEC
strains, which represents a feature of pathoadaptive changes in several bacterial pathogens
(50). The accumulation of these mutations confers AIEC the ability to adhere to CEACAM-
expressing intestinal epithelial cells and to participate in the development of chronic
inflammation in a genetically susceptible host. Finally, it is known that the long polar
fimbriae help AIEC to interact with intestinal Peyer’s patches and M cells (51). A recent
study analyzed the effect of gastrointestinal conditions on AIEC long polar fimbriae
expression. The authors found that bile salts strongly enhanced fimbriae expression, causing
a higher level of interaction of AIEC with Peyer’s patches and a higher level of translocation
through M cell monolayers (52).
Kalita et al. Page 5
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Regarding the clinical implications of AIEC recent investigation indicates that colonization
of AIEC results in chronic colitis in mice lacking the flagellin receptor TLR5. Transient
AIEC colonization drove intestinal inflammation which is associated with altering
microbiota composition (53). Proteases and protease inhibitors control microbiota
composition, immune response and intestinal function to maintain gut homeostasis. CYLD
is a de-ubiquitinase that is significantly downregulated in the intestine of Crohn’s disease
patients (54). Decrease CYLD expression results in an enhanced intracellular replication of
AIEC (54). Therefore, protection against AIEC during microbiota acquisition might be a
strategy to control IBD in genetically susceptible individuals (53).
Uropathogenic E. coli (UPEC)
UPEC are among the most prevalent extra-intestinal bacteria, accounting for 90% of all
urinary tract infection (UTI) (2). The most predominant chaperone-usher fimbriae in UPEC
strains is the type 1 fimbriae, which is an important determinant for pathogenicity, allowing
the interaction of UPEC with urinary tract host epithelia (55, 56). FimH within the type 1
fimbriae bound to α-D-mannosylated uroplakin, facilitating bacterial invasion, colonization
and formation of biofilm-like structures called intracellular bacterial communities (IBCs)
(57) (Figure 1). A high-throughput insilico analysis and in-vitro binding study discovered
that pathoadaptive alleles of FimH, with variant residues outside the binding pocket, affect
FimH-mediated acute and chronic pathogenesis of two prototype UPEC strains (56). The
study argues that FimH variants, which maintain a high-affinity conformation, were
attenuated during chronic bladder infection, implying FimH's ability to switch between
conformations is important during pathogenesis.
With respect to the regulatory mechanisms controlling UPEC colonization, Mitra et al found
UvrY as a key regulator modulating phase variation during UPEC pathogenesis, down-
regulating the expression of type 1 fimbrial structural genes, and influencing biofilm
formation, virulence and motility in UPEC strain CFT073 (58). Cpx is another key regulator
involved in bacterial adhesion. The deletion of cpxRA impaired the ability of UPEC strain
UTI89 to invade and colonize bladder epithelial cells, suggesting that the Cpx system is
needed for UPEC persistence in the urinary tract (59). Similarly, mutation in cpxRA and
cpxP in CFT073 also greatly reduced virulence tested the zebrafish infection model (59).
Finally, natural medicinal plants and secondary metabolites have being study because asiatic
acid and ursolic acid decreased expression of P fimbriae and curli fibers, altering cell
morphology and adhesion of UPEC to uroepithelial cells (60). Finally, Rafsanjany et al also
demonstrated anti-adhesive effects of various medicinal plant extracts against UPEC strains
(61).
Conclusion
The recent progress understanding the adhesion/invasion properties of intestinal and extra-
intestinal pathogenic E. coli during colonization of host cells reveals that there is a need of
further mechanistic studies that can be used for development of specific therapeutic
approaches.
Kalita et al. Page 6
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
of AIEC results in chronic colitis in mice lacking the flagellin receptor TLR5. Transient
AIEC colonization drove intestinal inflammation which is associated with altering
microbiota composition (53). Proteases and protease inhibitors control microbiota
composition, immune response and intestinal function to maintain gut homeostasis. CYLD
is a de-ubiquitinase that is significantly downregulated in the intestine of Crohn’s disease
patients (54). Decrease CYLD expression results in an enhanced intracellular replication of
AIEC (54). Therefore, protection against AIEC during microbiota acquisition might be a
strategy to control IBD in genetically susceptible individuals (53).
Uropathogenic E. coli (UPEC)
UPEC are among the most prevalent extra-intestinal bacteria, accounting for 90% of all
urinary tract infection (UTI) (2). The most predominant chaperone-usher fimbriae in UPEC
strains is the type 1 fimbriae, which is an important determinant for pathogenicity, allowing
the interaction of UPEC with urinary tract host epithelia (55, 56). FimH within the type 1
fimbriae bound to α-D-mannosylated uroplakin, facilitating bacterial invasion, colonization
and formation of biofilm-like structures called intracellular bacterial communities (IBCs)
(57) (Figure 1). A high-throughput insilico analysis and in-vitro binding study discovered
that pathoadaptive alleles of FimH, with variant residues outside the binding pocket, affect
FimH-mediated acute and chronic pathogenesis of two prototype UPEC strains (56). The
study argues that FimH variants, which maintain a high-affinity conformation, were
attenuated during chronic bladder infection, implying FimH's ability to switch between
conformations is important during pathogenesis.
With respect to the regulatory mechanisms controlling UPEC colonization, Mitra et al found
UvrY as a key regulator modulating phase variation during UPEC pathogenesis, down-
regulating the expression of type 1 fimbrial structural genes, and influencing biofilm
formation, virulence and motility in UPEC strain CFT073 (58). Cpx is another key regulator
involved in bacterial adhesion. The deletion of cpxRA impaired the ability of UPEC strain
UTI89 to invade and colonize bladder epithelial cells, suggesting that the Cpx system is
needed for UPEC persistence in the urinary tract (59). Similarly, mutation in cpxRA and
cpxP in CFT073 also greatly reduced virulence tested the zebrafish infection model (59).
Finally, natural medicinal plants and secondary metabolites have being study because asiatic
acid and ursolic acid decreased expression of P fimbriae and curli fibers, altering cell
morphology and adhesion of UPEC to uroepithelial cells (60). Finally, Rafsanjany et al also
demonstrated anti-adhesive effects of various medicinal plant extracts against UPEC strains
(61).
Conclusion
The recent progress understanding the adhesion/invasion properties of intestinal and extra-
intestinal pathogenic E. coli during colonization of host cells reveals that there is a need of
further mechanistic studies that can be used for development of specific therapeutic
approaches.
Kalita et al. Page 6
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
⊘ This is a preview!⊘
Do you want full access?
Subscribe today to unlock all pages.
Trusted by 1+ million students worldwide

Acknowledgments
The laboratory of A.G.T. was supported in part by NIH/NIAID grant AI079154.
References and Recommended reading
1. Leimbach A, Hacker J, Dobrindt U. E. coli as an all-rounder: the thin line between commensalism
and pathogenicity. Curr Top Microbiol Immunol. 2013; 358:3–32. [PubMed: 23340801]
2. Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nature Rev Microbiol. 2004; 2:123–
140. [PubMed: 15040260]
3. Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent advances in
understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev. 2013; 4:822–880. [PubMed:
24092857] . This is a comprehensive review highlighting recent advances in the understanding of
pathogenesis of intestinal pathotypes of E. coli.
4. Farfan MJ, Torres AG. Molecular mechanisms that mediate colonization of Shiga toxin-producing
Escherichia coli strains. Infect Immun. 2012; 80:903–913. [PubMed: 22144484]
5. Ferens WA, Hovde CJ. Escherichia coli O157:H7: animal reservoir and sources of human infection.
Foodborne Pathog Dis. 2011; 8:465–487. [PubMed: 21117940]
6. Zumbrun SD, Melton-Celsa AR, Smith MA, Gilbreath JJ, Merrell DS, O'Brien AD. Dietary choice
affects Shiga toxin-producing Escherichia coli (STEC) O157:H7 colonization and disease. Proc
Natl Acad Sci U S A. 2013; 110:E2126–E2133. [PubMed: 23690602] . This paper demonstrate that
susceptibility to infection and subsequent disease after E. coli O157:H7 consumption may depend,
on individual diet and/or commensal flora properties.
7. Sheng H, Nguyen YN, Hovde CJ, Sperandio V. SdiA aids enterohemorrhagic Escherichia coli
carriage by cattle fed a forage or grain diet. Infect Immun. 2013; 81:3472–3478. [PubMed:
23836826]
8. Hughes DT, Terekhova DA, Liou L, Hovde CJ, Sahl JW, Patankar AV, et al. Chemical sensing in
mammalian host-bacterial commensal associations. Proc Natl Acad Sci U S A. 2010; 107:9831–
9836. [PubMed: 20457895]
9. Garlanda C, Riva F, Polentarutti N, Buracchi C, Sironi M, De Bortoli M, et al. Intestinal
inflammation in mice deficient in Tir8, an inhibitory member of the IL-1 receptor family. Proc Natl
Acad Sci U S A. 2004; 101:3522–3526. [PubMed: 14993616]
10. Wald D, Qin J, Zhao Z, Qian Y, Naramura M, Tian L, et al. SIGIRR, a negative regulator of Toll-
like receptor-interleukin 1 receptor signaling. Nat Immunol. 2003; 4:920–927. [PubMed:
12925853]
11. Sham HP, Yu EY, Gulen MF, Bhinder G, Stahl M, Chan JM, et al. SIGIRR, a negative regulator of
TLR/IL-1R signalling promotes Microbiota dependent resistance to colonization by enteric
bacterial pathogens. PLoS Pathog. 2013; 9:e1003539. [PubMed: 23950714] . The paper
demonstrates that SIGIRR expression by intestinal epithelial cells is a strategy that promotes
commensal microbe-based colonization resistance against bacterial pathogens.
12. Hansen AM, Chaerkady R, Sharma J, Diaz-Mejia JJ, Tyagi N, Renuse S, et al. The Escherichia
coli phosphotyrosine proteome relates to core pathways and virulence. PLoS Pathog. 2013;
9:e1003403. [PubMed: 23785281]
13. Hansen AM, Jin DJ. SspA up-regulates gene expression of the LEE pathogenicity island by
decreasing H-NS levels in enterohemorrhagic Escherichia coli. BMC Microbiol. 2012; 12:231.
[PubMed: 23051860]
14. Branchu P, Matrat S, Vareille M, Garrivier A, Durand A, Crepin S, et al. NsrR, GadE, and GadX
interplay in repressing expression of the Escherichia coli O157:H7 LEE pathogenicity island in
response to nitric oxide. PLoS Pathog. 2014; 10:e1003874. [PubMed: 24415940]
15. Ochoa TJ, Barletta F, Contreras C, Mercado E. New insights into the epidemiology of
enteropathogenic Escherichia coli infection. Trans R Soc Trop Med Hyg. 2008; 102:852–856.
[PubMed: 18455741]
Kalita et al. Page 7
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
The laboratory of A.G.T. was supported in part by NIH/NIAID grant AI079154.
References and Recommended reading
1. Leimbach A, Hacker J, Dobrindt U. E. coli as an all-rounder: the thin line between commensalism
and pathogenicity. Curr Top Microbiol Immunol. 2013; 358:3–32. [PubMed: 23340801]
2. Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nature Rev Microbiol. 2004; 2:123–
140. [PubMed: 15040260]
3. Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent advances in
understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev. 2013; 4:822–880. [PubMed:
24092857] . This is a comprehensive review highlighting recent advances in the understanding of
pathogenesis of intestinal pathotypes of E. coli.
4. Farfan MJ, Torres AG. Molecular mechanisms that mediate colonization of Shiga toxin-producing
Escherichia coli strains. Infect Immun. 2012; 80:903–913. [PubMed: 22144484]
5. Ferens WA, Hovde CJ. Escherichia coli O157:H7: animal reservoir and sources of human infection.
Foodborne Pathog Dis. 2011; 8:465–487. [PubMed: 21117940]
6. Zumbrun SD, Melton-Celsa AR, Smith MA, Gilbreath JJ, Merrell DS, O'Brien AD. Dietary choice
affects Shiga toxin-producing Escherichia coli (STEC) O157:H7 colonization and disease. Proc
Natl Acad Sci U S A. 2013; 110:E2126–E2133. [PubMed: 23690602] . This paper demonstrate that
susceptibility to infection and subsequent disease after E. coli O157:H7 consumption may depend,
on individual diet and/or commensal flora properties.
7. Sheng H, Nguyen YN, Hovde CJ, Sperandio V. SdiA aids enterohemorrhagic Escherichia coli
carriage by cattle fed a forage or grain diet. Infect Immun. 2013; 81:3472–3478. [PubMed:
23836826]
8. Hughes DT, Terekhova DA, Liou L, Hovde CJ, Sahl JW, Patankar AV, et al. Chemical sensing in
mammalian host-bacterial commensal associations. Proc Natl Acad Sci U S A. 2010; 107:9831–
9836. [PubMed: 20457895]
9. Garlanda C, Riva F, Polentarutti N, Buracchi C, Sironi M, De Bortoli M, et al. Intestinal
inflammation in mice deficient in Tir8, an inhibitory member of the IL-1 receptor family. Proc Natl
Acad Sci U S A. 2004; 101:3522–3526. [PubMed: 14993616]
10. Wald D, Qin J, Zhao Z, Qian Y, Naramura M, Tian L, et al. SIGIRR, a negative regulator of Toll-
like receptor-interleukin 1 receptor signaling. Nat Immunol. 2003; 4:920–927. [PubMed:
12925853]
11. Sham HP, Yu EY, Gulen MF, Bhinder G, Stahl M, Chan JM, et al. SIGIRR, a negative regulator of
TLR/IL-1R signalling promotes Microbiota dependent resistance to colonization by enteric
bacterial pathogens. PLoS Pathog. 2013; 9:e1003539. [PubMed: 23950714] . The paper
demonstrates that SIGIRR expression by intestinal epithelial cells is a strategy that promotes
commensal microbe-based colonization resistance against bacterial pathogens.
12. Hansen AM, Chaerkady R, Sharma J, Diaz-Mejia JJ, Tyagi N, Renuse S, et al. The Escherichia
coli phosphotyrosine proteome relates to core pathways and virulence. PLoS Pathog. 2013;
9:e1003403. [PubMed: 23785281]
13. Hansen AM, Jin DJ. SspA up-regulates gene expression of the LEE pathogenicity island by
decreasing H-NS levels in enterohemorrhagic Escherichia coli. BMC Microbiol. 2012; 12:231.
[PubMed: 23051860]
14. Branchu P, Matrat S, Vareille M, Garrivier A, Durand A, Crepin S, et al. NsrR, GadE, and GadX
interplay in repressing expression of the Escherichia coli O157:H7 LEE pathogenicity island in
response to nitric oxide. PLoS Pathog. 2014; 10:e1003874. [PubMed: 24415940]
15. Ochoa TJ, Barletta F, Contreras C, Mercado E. New insights into the epidemiology of
enteropathogenic Escherichia coli infection. Trans R Soc Trop Med Hyg. 2008; 102:852–856.
[PubMed: 18455741]
Kalita et al. Page 7
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

16. Enserink R, Scholts R, Bruijning-Verhagen P, Duizer E, Vennema H, de Boer R, et al. High
detection rates of enteropathogens in asymptomatic children attending day care. PLoS ONE. 2014;
9:e89496. [PubMed: 24586825]
17. Yatsuyanagi J, Saito S, Sato H, Miyajima Y, Amano K, Enomoto K. Characterization of
enteropathogenic and enteroaggregative Escherichia coli isolated from diarrheal outbreaks. J Clin
Microbiol. 2002; 40:294–297. [PubMed: 11773137]
18. Hernandes RT, Elias WP, Vieira MA, Gomes TA. An overview of atypical enteropathogenic
Escherichia coli. FEMS Microbiol Lett. 2009; 297:137–149. [PubMed: 19527295]
19. Hernandes RT, De la Cruz MA, Yamamoto D, Giron JA, Gomes TA. Dissection of the role of pili
and type 2 and 3 secretion systems in adherence and biofilm formation of an atypical
enteropathogenic Escherichia coli strain. Infect Immun. 2013; 81:3793–3802. [PubMed:
23897608]
20. Kelly M, Hart E, Mundy R, Marches O, Wiles S, Badea L, et al. Essential role of the type III
secretion system effector NleB in colonization of mice by Citrobacter rodentium. Infect Immun.
2006; 74:2328–2337. [PubMed: 16552063]
21. Newton HJ, Pearson JS, Badea L, Kelly M, Lucas M, Holloway G, et al. The type III effectors
NleE and NleB from enteropathogenic E. coli and OspZ from Shigella block nuclear translocation
of NF-kappaB p65. PLoS Pathog. 2010; 6:e1000898. [PubMed: 20485572]
22. Gao X, Wang X, Pham TH, Feuerbacher LA, Lubos ML, Huang M, et al. NleB, a bacterial effector
with glycosyltransferase activity, targets GAPDH function to inhibit NF-kappaB activation. Cell
Host Microbe. 2013; 13:87–99. [PubMed: 23332158] . This paper reveals a virulence strategy
employed by Attaching and Effacing pathogens to inhibit NF-κB-dependent host innate immune
responses.
23. Pearson JS, Giogha C, Ong SY, Kennedy CL, Kelly M, Robinson KS, et al. A type III effector
antagonizes death receptor signalling during bacterial gut infection. Nature. 2013; 501:247–251.
[PubMed: 24025841]
24. Li S, Zhang L, Yao Q, Li L, Dong N, Rong J, et al. Pathogen blocks host death receptor signalling
by arginine GlcNAcylation of death domains. Nature. 2013; 501:242–246. [PubMed: 23955153] .
The paper reveals the mechanism of action of NleB, representing a new model by which bacteria
counteract host defenses.
25. Pan Q, Tian Y, Li X, Ye J, Liu Y, Song L, et al. Enhanced membrane-tethered mucin 3 (MUC3)
expression by a tetrameric branched peptide with a conserved TFLK motif inhibits bacteria
adherence. J Biol Chem. 2013; 288:5407–5416. [PubMed: 23316049]
26. Salcedo J, Barbera R, Matencio E, Alegria A, Lagarda MJ. Gangliosides and sialic acid effects
upon newborn pathogenic bacteria adhesion: an in vitro study. Food Chem. 2013; 136:726–734.
[PubMed: 23122120]
27. Beatty ME, Adcock PM, Smith SW, Quinlan K, Kamimoto LA, Rowe SY, et al. Epidemic diarrhea
due to enterotoxigenic Escherichia coli. Clin Infect Dis. 2006; 42:329–334. [PubMed: 16392076]
28. Torres AG, Zhou X, Kaper JB. Adherence of diarrheagenic Escherichia coli strains to epithelial
cells. Infect Immun. 2005; 73:18–29. [PubMed: 15618137]
29. Guevara CP, Luiz WB, Sierra A, Cruz C, Qadri F, Kaushik RS, et al. Enterotoxigenic Escherichia
coli CS21 pilus contributes to adhesion to intestinal cells and to pathogenesis under in vivo
conditions. Microbiology. 2013; 159:1725–1735. [PubMed: 23760820]
30. Fekete PZ, Mateo KS, Zhang W, Moxley RA, Kaushik RS, Francis DH. Both enzymatic and non-
enzymatic properties of heat-labile enterotoxin are responsible for LT-enhanced adherence of
enterotoxigenic Escherichia coli to porcine IPEC-J2 cells. Vet Microbiol. 2013; 164:330–335.
[PubMed: 23517763]
31. Kansal R, Rasko DA, Sahl JW, Munson GP, Roy K, Luo Q, et al. Transcriptional modulation of
enterotoxigenic Escherichia coli virulence genes in response to epithelial cell interactions. Infect
Immun. 2013; 81:259–270. [PubMed: 23115039] . This manuscript demonstrated that pathogen-
host interactions are finely coordinated by ETEC during the infectious process.
32. Zhou M, Duan Q, Zhu X, Guo Z, Li Y, Hardwidge PR, et al. Both flagella and F4 fimbriae from
F4ac+ enterotoxigenic Escherichia coli contribute to attachment to IPEC-J2 cells in vitro. Vet Res.
2013; 44:30. [PubMed: 23668601]
Kalita et al. Page 8
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
detection rates of enteropathogens in asymptomatic children attending day care. PLoS ONE. 2014;
9:e89496. [PubMed: 24586825]
17. Yatsuyanagi J, Saito S, Sato H, Miyajima Y, Amano K, Enomoto K. Characterization of
enteropathogenic and enteroaggregative Escherichia coli isolated from diarrheal outbreaks. J Clin
Microbiol. 2002; 40:294–297. [PubMed: 11773137]
18. Hernandes RT, Elias WP, Vieira MA, Gomes TA. An overview of atypical enteropathogenic
Escherichia coli. FEMS Microbiol Lett. 2009; 297:137–149. [PubMed: 19527295]
19. Hernandes RT, De la Cruz MA, Yamamoto D, Giron JA, Gomes TA. Dissection of the role of pili
and type 2 and 3 secretion systems in adherence and biofilm formation of an atypical
enteropathogenic Escherichia coli strain. Infect Immun. 2013; 81:3793–3802. [PubMed:
23897608]
20. Kelly M, Hart E, Mundy R, Marches O, Wiles S, Badea L, et al. Essential role of the type III
secretion system effector NleB in colonization of mice by Citrobacter rodentium. Infect Immun.
2006; 74:2328–2337. [PubMed: 16552063]
21. Newton HJ, Pearson JS, Badea L, Kelly M, Lucas M, Holloway G, et al. The type III effectors
NleE and NleB from enteropathogenic E. coli and OspZ from Shigella block nuclear translocation
of NF-kappaB p65. PLoS Pathog. 2010; 6:e1000898. [PubMed: 20485572]
22. Gao X, Wang X, Pham TH, Feuerbacher LA, Lubos ML, Huang M, et al. NleB, a bacterial effector
with glycosyltransferase activity, targets GAPDH function to inhibit NF-kappaB activation. Cell
Host Microbe. 2013; 13:87–99. [PubMed: 23332158] . This paper reveals a virulence strategy
employed by Attaching and Effacing pathogens to inhibit NF-κB-dependent host innate immune
responses.
23. Pearson JS, Giogha C, Ong SY, Kennedy CL, Kelly M, Robinson KS, et al. A type III effector
antagonizes death receptor signalling during bacterial gut infection. Nature. 2013; 501:247–251.
[PubMed: 24025841]
24. Li S, Zhang L, Yao Q, Li L, Dong N, Rong J, et al. Pathogen blocks host death receptor signalling
by arginine GlcNAcylation of death domains. Nature. 2013; 501:242–246. [PubMed: 23955153] .
The paper reveals the mechanism of action of NleB, representing a new model by which bacteria
counteract host defenses.
25. Pan Q, Tian Y, Li X, Ye J, Liu Y, Song L, et al. Enhanced membrane-tethered mucin 3 (MUC3)
expression by a tetrameric branched peptide with a conserved TFLK motif inhibits bacteria
adherence. J Biol Chem. 2013; 288:5407–5416. [PubMed: 23316049]
26. Salcedo J, Barbera R, Matencio E, Alegria A, Lagarda MJ. Gangliosides and sialic acid effects
upon newborn pathogenic bacteria adhesion: an in vitro study. Food Chem. 2013; 136:726–734.
[PubMed: 23122120]
27. Beatty ME, Adcock PM, Smith SW, Quinlan K, Kamimoto LA, Rowe SY, et al. Epidemic diarrhea
due to enterotoxigenic Escherichia coli. Clin Infect Dis. 2006; 42:329–334. [PubMed: 16392076]
28. Torres AG, Zhou X, Kaper JB. Adherence of diarrheagenic Escherichia coli strains to epithelial
cells. Infect Immun. 2005; 73:18–29. [PubMed: 15618137]
29. Guevara CP, Luiz WB, Sierra A, Cruz C, Qadri F, Kaushik RS, et al. Enterotoxigenic Escherichia
coli CS21 pilus contributes to adhesion to intestinal cells and to pathogenesis under in vivo
conditions. Microbiology. 2013; 159:1725–1735. [PubMed: 23760820]
30. Fekete PZ, Mateo KS, Zhang W, Moxley RA, Kaushik RS, Francis DH. Both enzymatic and non-
enzymatic properties of heat-labile enterotoxin are responsible for LT-enhanced adherence of
enterotoxigenic Escherichia coli to porcine IPEC-J2 cells. Vet Microbiol. 2013; 164:330–335.
[PubMed: 23517763]
31. Kansal R, Rasko DA, Sahl JW, Munson GP, Roy K, Luo Q, et al. Transcriptional modulation of
enterotoxigenic Escherichia coli virulence genes in response to epithelial cell interactions. Infect
Immun. 2013; 81:259–270. [PubMed: 23115039] . This manuscript demonstrated that pathogen-
host interactions are finely coordinated by ETEC during the infectious process.
32. Zhou M, Duan Q, Zhu X, Guo Z, Li Y, Hardwidge PR, et al. Both flagella and F4 fimbriae from
F4ac+ enterotoxigenic Escherichia coli contribute to attachment to IPEC-J2 cells in vitro. Vet Res.
2013; 44:30. [PubMed: 23668601]
Kalita et al. Page 8
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

33. Gonzalez-Ortiz G, Perez JF, Hermes RG, Molist F, Jimenez-Diaz R, Martin-Orue SM. Screening
the ability of natural feed ingredients to interfere with the adherence of enterotoxigenic
Escherichia coli (ETEC) K88 to the porcine intestinal mucus. Br J Nutr. 2014; 111:633–642.
[PubMed: 24047890]
34. Roberts CL, Keita AV, Parsons BN, Prorok-Hamon M, Knight P, Winstanley C, et al. Soluble
plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J Nutr Biochem.
2013; 24:97–103. [PubMed: 22818716]
35. Flores J, Okhuysen PC. Enteroaggregative Escherichia coli infection. Curr Opin Gastroenterol.
2009; 25:8–11. [PubMed: 19114769]
36. Weintraub A. Enteroaggregative Escherichia coli : epidemiology, virulence and detection. J Med
Microbiol. 2007; 56:4–8. [PubMed: 17172509]
37. Lima IF, Boisen N, Quetz Jda S, Havt A, de Carvalho EB, Soares AM, et al. Prevalence of
enteroaggregative Escherichia coli and its virulence-related genes in a case-control study among
children from north-eastern Brazil. J Med Microbiol. 2013; 62:683–693. [PubMed: 23429698]
38. Nuesch-Inderbinen MT, Hofer E, Hachler H, Beutin L, Stephan R. Characteristics of
enteroaggregative Escherichia coli isolated from healthy carriers and from patients with diarrhoea.
J Med Microbiol. 2013; 62:1828–1834. [PubMed: 24008499]
39. Munera D, Ritchie JM, Hatzios SK, Bronson R, Fang G, Schadt EE, et al. Autotransporters but not
pAA are critical for rabbit colonization by Shiga toxin-producing Escherichia coli O104:H4. Nat
Commun. 2014; 5:3080. [PubMed: 24445323] . This study revealed that the virulence pAA
plasmid in STEC O104:H4 is dispensable for intestinal colonization while the production of
autotransporters is critical for development of intestinal pathology.
40. Herzog K, Engeler Dusel J, Hugentobler M, Beutin L, Sagesser G, Stephan R, et al. Diarrheagenic
enteroaggregative Escherichia coli causing urinary tract infection and bacteremia leading to sepsis.
Infection. 2014; 42:441–444. [PubMed: 24323785]
41. Boll EJ, Struve C, Boisen N, Olesen B, Stahlhut SG, Krogfelt KA. Role of enteroaggregative
Escherichia coli virulence factors in uropathogenesis. Infect Immun. 2013; 81:1164–1171.
[PubMed: 23357383] . The study found that EAEC-specific virulence factors increase
uropathogenicity and strains carrying these factors might cause a community-acquired urinary
tract infections.
42. Jafari A, Shafaei E, Oloomi M, Aghasadeghi MR, Bouzari S. Genotypic and Phenotypic
Comparison of Enteroaggregative Escherichia coli Isolates from HIV-Positive and non-HIV
Diarrheal Samples. Curr HIV Res. 2014 [Epub ahead of print].
43. Cieza RJ, Cao AT, Cong Y, Torres AG. Immunomodulation for gastrointestinal infections. Expert
Rev Anti Infect Ther. 2012; 10:391–400. [PubMed: 22397571]
44. Rolhion N, Darfeuille-Michaud A. Adherent-invasive Escherichia coli in inflammatory bowel
disease. Bowel Dis. 2007; 13:1277–1283.
45. Glasser AL, Boudeau J, Barnich N, Perruchot MH, Colombel JF, Darfeuille-Michaud A. Adherent
invasive Escherichia coli strains from patients with Crohn's disease survive and replicate within
macrophages without inducing host cell death. Infect Immun. 2001; 69:5529–5537. [PubMed:
11500426]
46. Small CL, Reid-Yu SA, McPhee JB, Coombes BK. Persistent infection with Crohn's disease-
associated adherent-invasive Escherichia coli leads to chronic inflammation and intestinal fibrosis.
Nat Commun. 2013; 4:1957. [PubMed: 23748852] . This paper describes a murine model in which
chronic adherent-invasive E. coli infections result in immunopathology similar to that seen in
Crohn's disease patients.
47. Glas J, Seiderer J, Fries C, Tillack C, Pfennig S, Weidinger M, et al. CEACAM6 gene variants in
inflammatory bowel disease. PLoS One. 2011; 6:e19319. [PubMed: 21559399]
48. Barnich N, Carvalho FA, Glasser AL, Darcha C, Jantscheff P, Allez M, et al. CEACAM6 acts as a
receptor for adherent-invasive E. coli supporting ileal mucosa colonization in Crohn disease. J
Clin Invest. 2007; 117:1566–1574. [PubMed: 17525800]
49. Martinez-Medina M, Denizot J, Dreux N, Robin F, Billard E, Bonnet R, et al. Western diet induces
dysbiosis with increased E. coli in CEABAC10 mice, alters host barrier function favouring AIEC
colonisation. Gut. 2014; 63:116–124. [PubMed: 23598352]
Kalita et al. Page 9
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
the ability of natural feed ingredients to interfere with the adherence of enterotoxigenic
Escherichia coli (ETEC) K88 to the porcine intestinal mucus. Br J Nutr. 2014; 111:633–642.
[PubMed: 24047890]
34. Roberts CL, Keita AV, Parsons BN, Prorok-Hamon M, Knight P, Winstanley C, et al. Soluble
plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J Nutr Biochem.
2013; 24:97–103. [PubMed: 22818716]
35. Flores J, Okhuysen PC. Enteroaggregative Escherichia coli infection. Curr Opin Gastroenterol.
2009; 25:8–11. [PubMed: 19114769]
36. Weintraub A. Enteroaggregative Escherichia coli : epidemiology, virulence and detection. J Med
Microbiol. 2007; 56:4–8. [PubMed: 17172509]
37. Lima IF, Boisen N, Quetz Jda S, Havt A, de Carvalho EB, Soares AM, et al. Prevalence of
enteroaggregative Escherichia coli and its virulence-related genes in a case-control study among
children from north-eastern Brazil. J Med Microbiol. 2013; 62:683–693. [PubMed: 23429698]
38. Nuesch-Inderbinen MT, Hofer E, Hachler H, Beutin L, Stephan R. Characteristics of
enteroaggregative Escherichia coli isolated from healthy carriers and from patients with diarrhoea.
J Med Microbiol. 2013; 62:1828–1834. [PubMed: 24008499]
39. Munera D, Ritchie JM, Hatzios SK, Bronson R, Fang G, Schadt EE, et al. Autotransporters but not
pAA are critical for rabbit colonization by Shiga toxin-producing Escherichia coli O104:H4. Nat
Commun. 2014; 5:3080. [PubMed: 24445323] . This study revealed that the virulence pAA
plasmid in STEC O104:H4 is dispensable for intestinal colonization while the production of
autotransporters is critical for development of intestinal pathology.
40. Herzog K, Engeler Dusel J, Hugentobler M, Beutin L, Sagesser G, Stephan R, et al. Diarrheagenic
enteroaggregative Escherichia coli causing urinary tract infection and bacteremia leading to sepsis.
Infection. 2014; 42:441–444. [PubMed: 24323785]
41. Boll EJ, Struve C, Boisen N, Olesen B, Stahlhut SG, Krogfelt KA. Role of enteroaggregative
Escherichia coli virulence factors in uropathogenesis. Infect Immun. 2013; 81:1164–1171.
[PubMed: 23357383] . The study found that EAEC-specific virulence factors increase
uropathogenicity and strains carrying these factors might cause a community-acquired urinary
tract infections.
42. Jafari A, Shafaei E, Oloomi M, Aghasadeghi MR, Bouzari S. Genotypic and Phenotypic
Comparison of Enteroaggregative Escherichia coli Isolates from HIV-Positive and non-HIV
Diarrheal Samples. Curr HIV Res. 2014 [Epub ahead of print].
43. Cieza RJ, Cao AT, Cong Y, Torres AG. Immunomodulation for gastrointestinal infections. Expert
Rev Anti Infect Ther. 2012; 10:391–400. [PubMed: 22397571]
44. Rolhion N, Darfeuille-Michaud A. Adherent-invasive Escherichia coli in inflammatory bowel
disease. Bowel Dis. 2007; 13:1277–1283.
45. Glasser AL, Boudeau J, Barnich N, Perruchot MH, Colombel JF, Darfeuille-Michaud A. Adherent
invasive Escherichia coli strains from patients with Crohn's disease survive and replicate within
macrophages without inducing host cell death. Infect Immun. 2001; 69:5529–5537. [PubMed:
11500426]
46. Small CL, Reid-Yu SA, McPhee JB, Coombes BK. Persistent infection with Crohn's disease-
associated adherent-invasive Escherichia coli leads to chronic inflammation and intestinal fibrosis.
Nat Commun. 2013; 4:1957. [PubMed: 23748852] . This paper describes a murine model in which
chronic adherent-invasive E. coli infections result in immunopathology similar to that seen in
Crohn's disease patients.
47. Glas J, Seiderer J, Fries C, Tillack C, Pfennig S, Weidinger M, et al. CEACAM6 gene variants in
inflammatory bowel disease. PLoS One. 2011; 6:e19319. [PubMed: 21559399]
48. Barnich N, Carvalho FA, Glasser AL, Darcha C, Jantscheff P, Allez M, et al. CEACAM6 acts as a
receptor for adherent-invasive E. coli supporting ileal mucosa colonization in Crohn disease. J
Clin Invest. 2007; 117:1566–1574. [PubMed: 17525800]
49. Martinez-Medina M, Denizot J, Dreux N, Robin F, Billard E, Bonnet R, et al. Western diet induces
dysbiosis with increased E. coli in CEABAC10 mice, alters host barrier function favouring AIEC
colonisation. Gut. 2014; 63:116–124. [PubMed: 23598352]
Kalita et al. Page 9
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
⊘ This is a preview!⊘
Do you want full access?
Subscribe today to unlock all pages.
Trusted by 1+ million students worldwide

50. Dreux N, Denizot J, Martinez-Medina M, Mellmann A, Billig M, Kisiela D, et al. Point mutations
in FimH adhesin of Crohn's disease-associated adherent-invasive Escherichia coli enhance
intestinal inflammatory response. PLoS Pathog. 2013; 9:e1003141. [PubMed: 23358328] . This
study highlights a mechanism of AIEC virulence evolution that leads to the development of
chronic inflammatory bowel disease in a genetically susceptible host.
51. Chassaing B, Rolhion N, de Vallee A, Salim SY, Prorok-Hamon M, Neut C, et al. Crohn disease--
associated adherent-invasive E. coli bacteria target mouse and human Peyer's patches via long
polar fimbriae. J Clin Invest. 2011; 121:966–975. [PubMed: 21339647]
52. Chassaing B, Etienne-Mesmin L, Bonnet R, Darfeuille-Michaud A. Bile salts induce long polar
fimbriae expression favouring Crohn's disease-associated adherent-invasive Escherichia coli
interaction with Peyer's patches. Environ Microbiol. 2013; 15:355–371. [PubMed: 22789019]
53. Chassaing B, Koren O, Carvalho FA, Ley RE, Gewirtz AT. AIEC pathobiont instigates chronic
colitis in susceptible hosts by altering microbiota composition. Gut. 2014; 63:1069–1080.
[PubMed: 23896971]
54. Cleynen I, Vazeille E, Artieda M, Verspaget HW, Szczypiorska M, Bringer MA, et al. Genetic and
microbial factors modulating the ubiquitin proteasome system in inflammatory bowel disease. Gut.
2013 [Epub ahead of print].
55. Sauer FG, Mulvey MA, Schilling JD, Martinez JJ, Hultgren SJ. Bacterial pili: molecular
mechanisms of pathogenesis. Curr Opin Microbiol. 2000; 3:65–72. [PubMed: 10679419]
56. Schwartz DJ, Kalas V, Pinkner JS, Chen SL, Spaulding CN, Dodson KW, et al. Positively selected
FimH residues enhance virulence during urinary tract infection by altering FimH conformation.
Proc Natl Acad Sci U S A. 2013; 110:15530–15537. [PubMed: 24003161] . This study present
evidence indicating that positively selected residues within type 1 fimbriae modulate bacterial
fitness during UTI by affecting FimH conformation and function.
57. Anderson GG, Palermo JJ, Schilling JD, Roth R, Heuser J, Hultgren SJ. Intracellular bacterial
biofilm-like pods in urinary tract infections. Science. 2003; 301:105–107. [PubMed: 12843396]
58. Mitra A, Palaniyandi S, Herren CD, Zhu X, Mukhopadhyay S. Pleiotropic roles of uvrY on biofilm
formation, motility and virulence in uropathogenic Escherichia coli CFT073. PLoS One. 2013;
8:e55492. [PubMed: 23383333]
59. Debnath I, Norton JP, Barber AE, Ott EM, Dhakal BK, Kulesus RR, et al. The Cpx stress response
system potentiates the fitness and virulence of uropathogenic Escherichia coli. Infect Immun.
2013; 81:1450–1459. [PubMed: 23429541]
60. Dorota W, Marta K, Dorota TG. Effect of asiatic and ursolic acids on morphology, hydrophobicity,
and adhesion of UPECs to uroepithelial cells. Folia Microbiol (Praha). 2013; 58:245–252.
[PubMed: 23132656]
61. Rafsanjany N, Lechtenberg M, Petereit F, Hensel A. Antiadhesion as a functional concept for
protection against uropathogenic Escherichia coli : in vitro studies with traditionally used plants
with antiadhesive activity against uropathognic Escherichia coli. J Ethnopharmacol. 2013;
145:591–597. [PubMed: 23211661]
Kalita et al. Page 10
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
in FimH adhesin of Crohn's disease-associated adherent-invasive Escherichia coli enhance
intestinal inflammatory response. PLoS Pathog. 2013; 9:e1003141. [PubMed: 23358328] . This
study highlights a mechanism of AIEC virulence evolution that leads to the development of
chronic inflammatory bowel disease in a genetically susceptible host.
51. Chassaing B, Rolhion N, de Vallee A, Salim SY, Prorok-Hamon M, Neut C, et al. Crohn disease--
associated adherent-invasive E. coli bacteria target mouse and human Peyer's patches via long
polar fimbriae. J Clin Invest. 2011; 121:966–975. [PubMed: 21339647]
52. Chassaing B, Etienne-Mesmin L, Bonnet R, Darfeuille-Michaud A. Bile salts induce long polar
fimbriae expression favouring Crohn's disease-associated adherent-invasive Escherichia coli
interaction with Peyer's patches. Environ Microbiol. 2013; 15:355–371. [PubMed: 22789019]
53. Chassaing B, Koren O, Carvalho FA, Ley RE, Gewirtz AT. AIEC pathobiont instigates chronic
colitis in susceptible hosts by altering microbiota composition. Gut. 2014; 63:1069–1080.
[PubMed: 23896971]
54. Cleynen I, Vazeille E, Artieda M, Verspaget HW, Szczypiorska M, Bringer MA, et al. Genetic and
microbial factors modulating the ubiquitin proteasome system in inflammatory bowel disease. Gut.
2013 [Epub ahead of print].
55. Sauer FG, Mulvey MA, Schilling JD, Martinez JJ, Hultgren SJ. Bacterial pili: molecular
mechanisms of pathogenesis. Curr Opin Microbiol. 2000; 3:65–72. [PubMed: 10679419]
56. Schwartz DJ, Kalas V, Pinkner JS, Chen SL, Spaulding CN, Dodson KW, et al. Positively selected
FimH residues enhance virulence during urinary tract infection by altering FimH conformation.
Proc Natl Acad Sci U S A. 2013; 110:15530–15537. [PubMed: 24003161] . This study present
evidence indicating that positively selected residues within type 1 fimbriae modulate bacterial
fitness during UTI by affecting FimH conformation and function.
57. Anderson GG, Palermo JJ, Schilling JD, Roth R, Heuser J, Hultgren SJ. Intracellular bacterial
biofilm-like pods in urinary tract infections. Science. 2003; 301:105–107. [PubMed: 12843396]
58. Mitra A, Palaniyandi S, Herren CD, Zhu X, Mukhopadhyay S. Pleiotropic roles of uvrY on biofilm
formation, motility and virulence in uropathogenic Escherichia coli CFT073. PLoS One. 2013;
8:e55492. [PubMed: 23383333]
59. Debnath I, Norton JP, Barber AE, Ott EM, Dhakal BK, Kulesus RR, et al. The Cpx stress response
system potentiates the fitness and virulence of uropathogenic Escherichia coli. Infect Immun.
2013; 81:1450–1459. [PubMed: 23429541]
60. Dorota W, Marta K, Dorota TG. Effect of asiatic and ursolic acids on morphology, hydrophobicity,
and adhesion of UPECs to uroepithelial cells. Folia Microbiol (Praha). 2013; 58:245–252.
[PubMed: 23132656]
61. Rafsanjany N, Lechtenberg M, Petereit F, Hensel A. Antiadhesion as a functional concept for
protection against uropathogenic Escherichia coli : in vitro studies with traditionally used plants
with antiadhesive activity against uropathognic Escherichia coli. J Ethnopharmacol. 2013;
145:591–597. [PubMed: 23211661]
Kalita et al. Page 10
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Paraphrase This Document
Need a fresh take? Get an instant paraphrase of this document with our AI Paraphraser

Key points
• Recent studies revealed how host signaling responses alter pathogenic E. coli
colonization.
• Advances in the understanding for the role of pathoadaptive mutations on
fimbrial adhesins and their contribution to the pathogenic process.
• Development of new murine model of chronic E. coli infection and the study of
novel E. coli pathotypes.
Kalita et al. Page 11
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
• Recent studies revealed how host signaling responses alter pathogenic E. coli
colonization.
• Advances in the understanding for the role of pathoadaptive mutations on
fimbrial adhesins and their contribution to the pathogenic process.
• Development of new murine model of chronic E. coli infection and the study of
novel E. coli pathotypes.
Kalita et al. Page 11
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
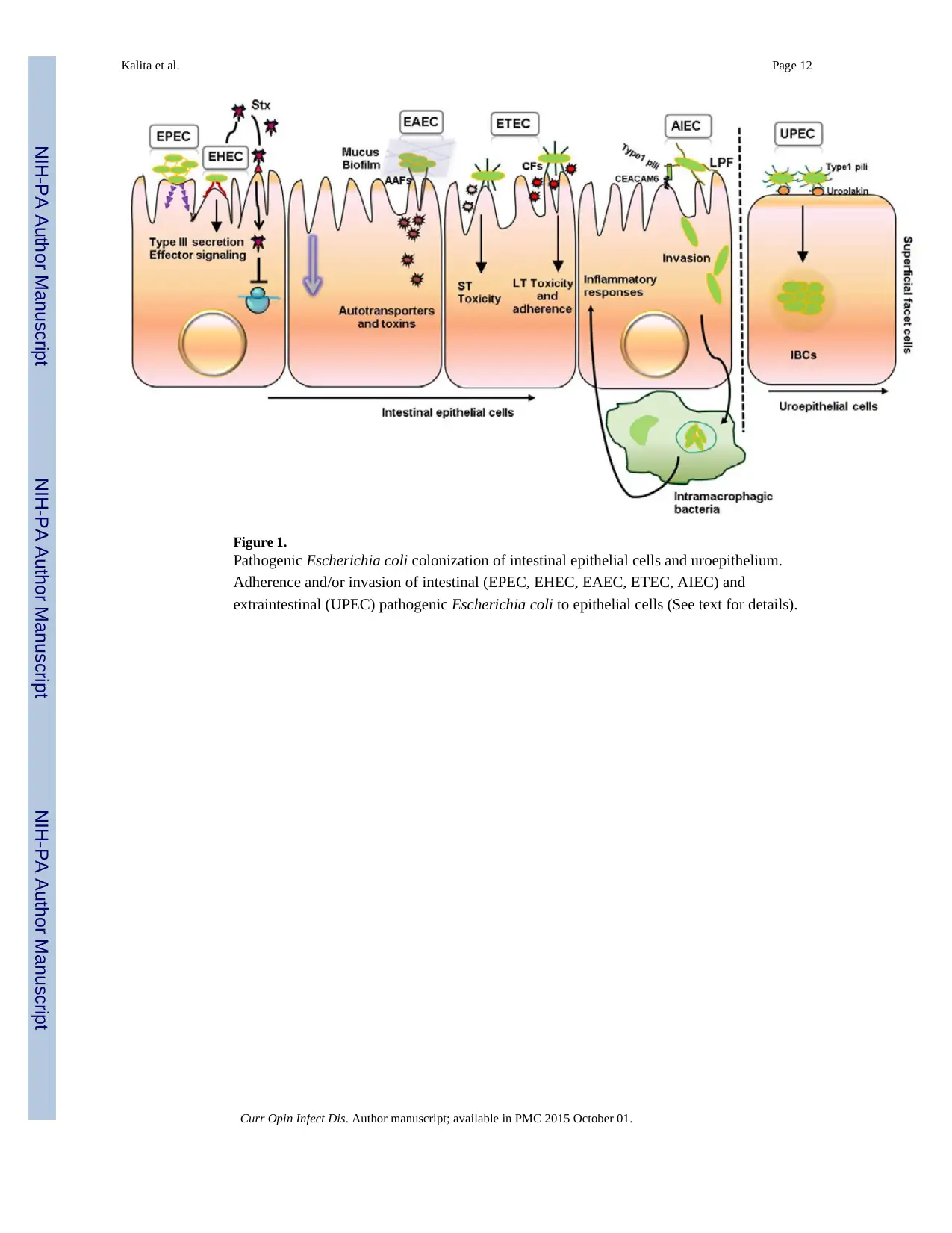
Figure 1.
Pathogenic Escherichia coli colonization of intestinal epithelial cells and uroepithelium.
Adherence and/or invasion of intestinal (EPEC, EHEC, EAEC, ETEC, AIEC) and
extraintestinal (UPEC) pathogenic Escherichia coli to epithelial cells (See text for details).
Kalita et al. Page 12
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Pathogenic Escherichia coli colonization of intestinal epithelial cells and uroepithelium.
Adherence and/or invasion of intestinal (EPEC, EHEC, EAEC, ETEC, AIEC) and
extraintestinal (UPEC) pathogenic Escherichia coli to epithelial cells (See text for details).
Kalita et al. Page 12
Curr Opin Infect Dis. Author manuscript; available in PMC 2015 October 01.
NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
⊘ This is a preview!⊘
Do you want full access?
Subscribe today to unlock all pages.
Trusted by 1+ million students worldwide
1 out of 12
Your All-in-One AI-Powered Toolkit for Academic Success.
+13062052269
info@desklib.com
Available 24*7 on WhatsApp / Email
![[object Object]](/_next/static/media/star-bottom.7253800d.svg)
Unlock your academic potential
Copyright © 2020–2026 A2Z Services. All Rights Reserved. Developed and managed by ZUCOL.