Report: Metabolic Engineering of Omega 3 Fatty Acids in E. coli
VerifiedAdded on 2023/04/24
|7
|1552
|100
Report
AI Summary
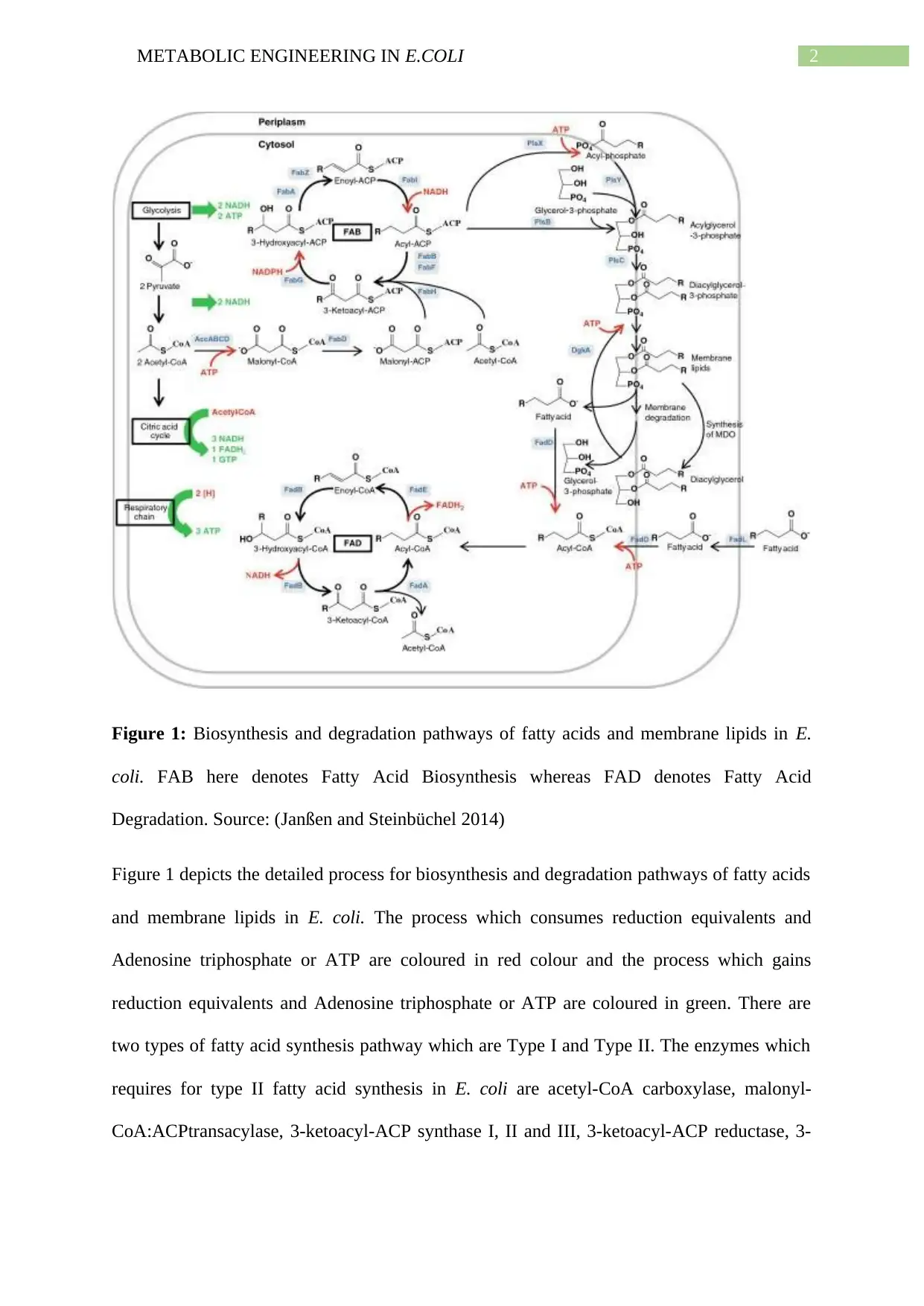
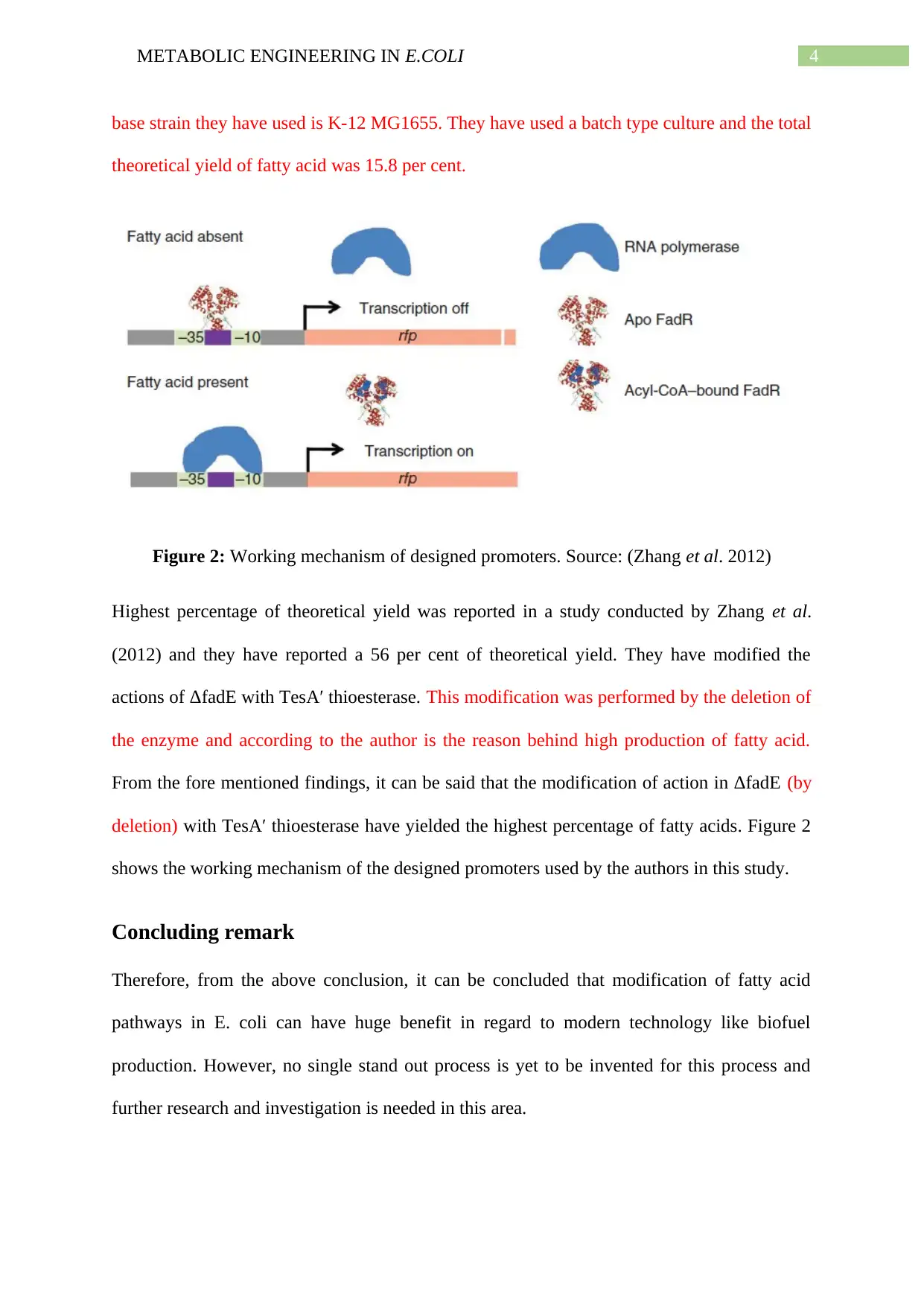
This report delves into the metabolic engineering of omega 3 fatty acids in E. coli, utilizing lignocellulosic hydrolysate as a base. It begins by outlining the normal pathway of fatty acid production in E. coli, highlighting the enzymes and processes involved, such as the formation of malonyl-CoA and the role of acyl carrier protein (ACP). The report then explores various modification strategies to enhance fatty acid synthesis, including increasing export rates, regulating membrane saturation, addressing metabolic bottlenecks, and conducting structural studies. It identifies the modification of ΔfadE with TesA′ thioesterase as a promising route, citing studies that have achieved significant theoretical yields of fatty acids through this method. The report concludes that while modifying fatty acid pathways in E. coli holds great potential for applications like biofuel production, further research is needed to optimize the process and achieve consistent, high yields. The report references several studies and includes figures illustrating biosynthesis and the working mechanism of designed promoters.





1 out of 7
Related Documents





Your All-in-One AI-Powered Toolkit for Academic Success.
+13062052269
info@desklib.com
Available 24*7 on WhatsApp / Email
![[object Object]](/_next/static/media/star-bottom.7253800d.svg)
Copyright © 2020–2026 A2Z Services. All Rights Reserved. Developed and managed by ZUCOL.